Филлотаксис

Филлотаксис (φύλλον с др.-греч. — «лист» и τάξις с др.-греч. — «расположение») или филлотаксия или листорасположение — это порядок расположения листьев на стебле растения.
По мере роста стебля листья располагаются на нём в определённом порядке, который обусловливает оптимальный доступ к свету. Листья появляются на стебле по спирали, как по часовой стрелке, так и против неё, под определённым углом расхождения (дивергенции). Филлотаксические спирали образуют особый класс узоров в природе.
История[править | править код]
Некоторые ранние ученые, особенно Леонардо да Винчи, наблюдали за спиральным расположением растений[1][2].
В 1754 году Шарль Бонне заметил, что спиральный филлотаксис растений часто выражается в рядах золотого сечения как по часовой, так и против часовой стрелки[3].
Математические наблюдения филлотаксиса последовали за работами немецких ботаников Карла Фридриха Шимпера и его друга Александра Брауна 1835 и 1830 годов соответственно; и почти одновременно французский ботаник Огюст Браве и его брат Луи связали коэффициенты филлотаксиса с последовательностью Фибоначчи в 1837 году[3].
Изучая и сравнивая характер расположения листьев на стебле у различных групп растений, Браун и Шимпер открыли, что при спиральном листорасположении, наиболее характерном для высших растений, места прикрепления листьев подчиняются математическим закономерностям, которые они выразили в виде дроби: числителем дроби является число оборотов спирали, проведенное на стебле между двумя листьями, расположенными друг над другом на одной прямой, так называемой ортоcтихе, а знаменателем — число листьев (листовой цикл) на этой спирали, не считая последнего листа. Однако объяснение найденной закономерности немецкими и французскими ос- нователями учения о филлотаксисе было идеалистическим и основывалось на «извечном» плане строения растений[4].

Понимание механизма пришлось ждать, пока Вильгельм Гофмейстер не предложил модель в 1868 году. Зачаток, зарождающийся лист, формируется в наименее переполненной части меристемы побега. Золотой угол между последовательными листьями — слепой результат этого толкания. Поскольку трех золотых дуг в сумме более чем достаточно, чтобы обернуть круг, это гарантирует, что никакие два листа никогда не будут следовать одной и той же радиальной линии от центра к краю. Генеративная спираль является следствием того же процесса, который производит спирали по часовой стрелке и против часовой стрелки, которые возникают в плотно упакованных структурах растений, таких как цветочные диски Protea или чешуи сосновых шишек.
В наше время такие исследователи, как Мэри Сноу и Джордж Сноу[5], продолжили эти исследования. Компьютерное моделирование и морфологические исследования подтвердили и уточнили идеи Хоффмайстера. Остаются вопросы по деталям. Ботаники расходятся во мнениях относительно того, зависит ли контроль миграции листьев от химических градиентов среди зачатков или от чисто механических сил. У некоторых растений наблюдались числа Люка, а не числа Фибоначчи, и иногда расположение листьев кажется случайным.
Основные типы филлотаксиса[править | править код]
Различают несколько основных вариантов листорасположения:
Очерёдное листорасположение[править | править код]
Очерёдное листорасположение (или последовательное, или рассеянное или спиральное) (лат. folia sparsa, f. alterna) — листья располагаются по одному (в очередь) на каждый узел[6]. Данный тип листоположения наиболее распространён. Примеры растений: берёза, яблоня, роза, традесканция, циссус, пеларгония.
При очерёдном листорасположении основания или вершины последовательных листьев можно соединить условной спиральной линией — основной генетической спиралью[7] (отражает последовательность заложения листьев — генезис).
-
 Очерёдное расположение листьев по спирали
Очерёдное расположение листьев по спирали -
 Спиральное расположение листьев у подсолнечника однолетнего (Helianthus annuus)
Спиральное расположение листьев у подсолнечника однолетнего (Helianthus annuus) -
 Aloe dorotheae со спиральным расположением листьев
Aloe dorotheae со спиральным расположением листьев -
 Aloe polyphylla со спиральным расположением листьев
Aloe polyphylla со спиральным расположением листьев
.svg)



Двурядное листорасположение[править | править код]
Двурядное или двустрочное листорасположение (лат. disticha)[8] — частный случай очерёдного листорасположения, когда угол расхождения равен 180° и все листья лежат в одной плоскости, то по одну, то по другую сторону от оси, отражает маятниковую симметрию деятельности апекса. Данный тип листорасположения характерен, например, для семейства Злаки (Poaceae) и рода Ситник (Juncus)[6]. Помимо злаковых встречается и у некоторых лилейных (гастерия), ирисовых (ирис, гладиолус), бобовых (горошек)[9].
-
 Двухрядное листорасположение — супротивное расположение листьев в одной плоскости
Двухрядное листорасположение — супротивное расположение листьев в одной плоскости -
 Равенала мадагаскарская (Ravenala madagascariensis) с двухрядным расположением листьев
Равенала мадагаскарская (Ravenala madagascariensis) с двухрядным расположением листьев -
 Кливия киноварная (Clivia miniata) с двухрядным расположением листьев
Кливия киноварная (Clivia miniata) с двухрядным расположением листьев -
 Боофона двурядная (Boophone disticha) с двухрядным расположением листьев
Боофона двурядная (Boophone disticha) с двухрядным расположением листьев -
 Gasteria obliqua с двухрядным расположением листьев
Gasteria obliqua с двухрядным расположением листьев -
 Ветвь тиса ягодного (Taxus baccata) с двурядным расположением игольчатых листьев
Ветвь тиса ягодного (Taxus baccata) с двурядным расположением игольчатых листьев -
 Aloe suprafoliata с двухрядным расположением листьев
Aloe suprafoliata с двухрядным расположением листьев
.svg)






Трёхрядное листорасположение[править | править код]
У Осоковых (Cyperaceae) листоположение трёхрядное[6], с углом расхождения 120°. Более сложные спирали см. ниже.
Однобочное листорасположение[править | править код]
Однобочное листорасположение (лат. folia unilateralia)[10] — частный случай очередного расположения, когда листья обращены в одну сторону. Угол расхождения составляет 360°.
-
 Однобочное расположение листьев
Однобочное расположение листьев -
 Купена аптечная (Polygonatum odoratum) c однобочным расположением листьев
Купена аптечная (Polygonatum odoratum) c однобочным расположением листьев -
![Костус прекрасный (Hellenia speciosa)[11] c однобочным расположением листьев](//upload.wikimedia.org/wikipedia/commons/thumb/a/a0/Starr-110215-1172-Costus_speciosus-leaves-KiHana_Nursery_Kihei-Maui_%2824448906923%29.jpg/120px-Starr-110215-1172-Costus_speciosus-leaves-KiHana_Nursery_Kihei-Maui_%2824448906923%29.jpg)
.svg)

![Костус прекрасный (Hellenia speciosa)[11] c однобочным расположением листьев](/wiki/%D0%A4%D0%B0%D0%B9%D0%BB:Starr-110215-1172-Costus_speciosus-leaves-KiHana_Nursery_Kihei-Maui_(24448906923).jpg)
Супротивное[править | править код]
Супроти́вное листорасположение (лат. folia opposita)[12][10] — листья располагаются по два на каждом узле. Пара противоположных листьев может рассматриваться как мутовка из двух листьев.
Попарно супротивное[править | править код]
Попарно супротивное или декуссатное листорасположение[13] (лат. folia opposita) — самый распространенный подвид супротивного листорасположения, каждый последующий узел на стебле повёрнут относительно предыдущего на угол 90°.
-
 Супротивное листорасположение с поворотом 90°, декуссатное расположение
Супротивное листорасположение с поворотом 90°, декуссатное расположение -
 Базилик тонкоцветный (Ocimum tenuiflorum) c декуссатным расположением листьев
Базилик тонкоцветный (Ocimum tenuiflorum) c декуссатным расположением листьев -
 Горечавка ластовневая (Gentiana asclepiadea) с супротивными листьями
Горечавка ластовневая (Gentiana asclepiadea) с супротивными листьями -
 Очиток видный (Sedum spectabile) c декуссатным расположением листьев
Очиток видный (Sedum spectabile) c декуссатным расположением листьев
.svg)
.jpg)


Двухрядное супротивное[править | править код]
Двухрядное супротивное листорасположение — частный случай супротивного листорасположения, когда все узлы лежат в одной плоскости. Примеры растений: сирень, яснотка, мята, жасмин, фуксия.
-
 Супротивное расположение листьев в одной плоскости
Супротивное расположение листьев в одной плоскости -
 Кизил обыкновенный (Cornus mas) с супротивными листьями, лежащими в одной плоскости
Кизил обыкновенный (Cornus mas) с супротивными листьями, лежащими в одной плоскости -
 Сирень Syringa × josiflexa с супротивными листьями, лежащими в одной плоскости
Сирень Syringa × josiflexa с супротивными листьями, лежащими в одной плоскости -

.svg)



Неравное супротивное[править | править код]
Неравное листорасположение (лат. folia disparia)[10] — супротивное расположение, когда два супротивных листа разные, как у Atropa belladonna.
-
 Неравное листорасположение листьев
Неравное листорасположение листьев -
 Красавка обыкновенная (Atropa belladonna) с неравным расположением листьев.
Красавка обыкновенная (Atropa belladonna) с неравным расположением листьев.


Мутовчатое[править | править код]
Муто́вчатое листорасположение (лат. folia verticillata) — листья располагаются по три и более на каждом узле стебля — мутовке. То есть, в каждом узле закладывается несколько листовых помордиев[7]. Большей частью число листьев в мутовке у каждого вида строго фиксировано, реже колеблется (от 2 до 4 y вербейника обыкновенного (Lysimachia vulgaris), дербенника иволистного (Lythrum salicaria))[6].
Как и супротивные листья, мутовки могут быть перекрёстными, когда каждая последующая мутовка повёрнута относительно предыдущей на угол 90°, или на половину угла между листьями. Супротивные листья могут показаться мутовчатыми на конце стебля (элодея, вороний глаз, олеандр).
Расположение мутовок довольно необычно для растений, за исключением растений с особенно короткими междоузлиями. Примерами деревьев с мутовчатым филлотаксисом являются Brabejum stellatifolium[14] и родственный род Макадамия (Macadamia)[15].
-
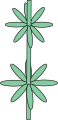 Мутовчатое листорасположение
Мутовчатое листорасположение -
 Хвостник обыкновенный (Hippuris vulgaris) с мутовчатым расположением листьев
Хвостник обыкновенный (Hippuris vulgaris) с мутовчатым расположением листьев -
 Вероникаструм виргинский (Veronicastrum virginicum) имеет по пять листьев в мутовках
Вероникаструм виргинский (Veronicastrum virginicum) имеет по пять листьев в мутовках -
 Расположение листьев у элодеи Нутталя (Elodea nuttallii) супротивное, но выглядит мутовчатым.
Расположение листьев у элодеи Нутталя (Elodea nuttallii) супротивное, но выглядит мутовчатым. -
 Myriophyllum quitense с мутовчатым расположением листьев
Myriophyllum quitense с мутовчатым расположением листьев
.svg)
.jpg)



Розеточное[править | править код]
Розе́точное листорасположение — листья, расположенные в розетке — прикорневой (базальной) мутовке с большим количеством разложенных по кругу листьев. Все листья находятся на одной высоте и расположены по кругу (камнеломка, хлорофитум, агава). Мутовка может возникать как базальная структура, в которой все листья прикреплены к основанию побега, а междоузлия маленькие или отсутствуют.

.jpg)
Прочие типы[править | править код]
Орикса японская (Orixa japonica) демонстрирует своеобразную филлотаксию, которая получила название «Тип Orixa». Последовательность листьев следующая: маленький лист на левой стороне, большой лист на левой стороне, маленький лист на правой стороне, большой лист на правой стороне и т. д., причём большие листья появляются больше на верхней стороне горизонтально растущих побегов, а маленькие листья — на нижней стороне[16].
-
 Орикса японская (Orixa japonica)
Орикса японская (Orixa japonica) -
 Орикса японская (Orixa japonica)
Орикса японская (Orixa japonica)


Закономерности листоположения[править | править код]
Основным правилом листорасположения является правило эквидистантности — равенства угловых расстояний между медианами смежных листьев. Из правила следуют некоторые следствия:
- Правило чередования кругов: листья новой мутовки закладываются точно в промежутках между листьями уже заложившейся мутовки.
- На оси побега обнаруживаются вертикальные прямые ряды листьев, расположенных точно друг над другом — ортостихи. На побеге с мутовчатым листорасположением их число, как правило, в два раза больше числа листьев в мутовке. Число листьев на отрезке генетической спирали, ограниченном двумя следующими друг за другом листьями одной ортостихи, называют листовым циклом[9].
В угле расхождения замечена точная последовательность Фибоначчи, выраженная дробью: 1/2, 1/3, 2/5, 3/8, 5/13, 8/21, 13/34, 21/55, 34/89. Дробь, выражающую значение угла дивергенции не в градусах, а в долях окружности, называют формулой листорасположения. Такая последовательность ограничена полным оборотом в 360°, 360° x 34/89 = 137,52 или 137° 30’28-29'' — угол, в математике известный под названием золотой угол[17][18], считается «идеальным» углом, при котором ни один лист на побеге, в теории, не может оказаться над другим; особенно встречается в расположении листовых зачатков на конусе нарастания в почке. Числитель в последовательности показывает количество оборотов спирали в пределах одного цикла. Первый лист следующего цикла будет располагаться над первым листом предыдущего. Знаменатель показывает число ортостих и, соответственно, листьев в одном листовом цикле.
Примеры формул листорасположения с соответствующими углами дивергенции:
- 1/2 (180°): два листа в обороте — двурядное листорасположение (злаки)
- 1/3 (120°): три листа в обороте — трёхрядное (семейство осоковых, лилейных, ольхи серой)
- 2/5 (144°): пять листьев за два оборота (дуб, лещина, табак, алоэ)
- 3/8 (135°): восемь листьев за три оборота (подорожник, капуста)
- 5/13 (138°27', молодило)
- 8/21 (137°8', лён)

Для удобства используют условные чертежи, изображающие проекцию всех узлов побега с их листьями на горизонтальную плоскость — диаграммы. Ось побега представлена в виде окружности, центр которой — его апекс. Последовательные узлы изображаются в виде концентрических окружностей или плоскостной спирали, условно соединяющей основания последовательных листьев. Листья обычно изображаются в виде фигурных скобок, символизирующих поперечный разрез листовой пластинки.
Порядок заложения листовых примордиев на апексе побега — наследственный признак, характерный для вида, рода или даже целого семейства растений, зависящий от генетических факторов. Кроме того, в процессе развёртывания побега из почки и его последующего роста на расположение листьев влияние оказывают и внешние факторы: условия освещения, сила тяжести, действие которой зависит от направления роста самого побега[9].
Контроль через гормоны[править | править код]
Первичный рост растения происходит в верхушечной меристеме (так называемая верхушка , также почка). Верхушка в процессе роста вращается вокруг своей оси; всегда образуются листовые зачатки, то есть ЧАС. Листовые почки, из которых впоследствии формируются листья.
Гормон ауксин транспортируется от верхушки листа к почке. Поглощение ауксина уже существующими зачатками направляет размещение нового зачатка. Ауксин поглощается уже существующими зачатками и, таким образом, удаляется из непосредственной близости (латеральная редукция). Новый зачаток не может развиваться непосредственно рядом со старым листовым зачатком, так как накопление ауксина может начаться только на определённом минимальном расстоянии. Поскольку самая молодая листовая почка поглощает ауксин больше, чем вторая по возрасту, новый зачаток возникает ближе ко второму самому молодому, чем к самому молодому зачатку. По этой причине угол расхождения двух последовательно образовавшихся зачатков соответствует типичному углу 137,5° («золотое сечение»).
В искусстве и архитектуре[править | править код]

Филлотаксис был использован в качестве источника вдохновения для ряда скульптур и архитектурных проектов. Акио Хизуме построил и продемонстрировал несколько бамбуковых башен, основанных на последовательности Фибоначчи, которые демонстрируют филлотаксис. Салех Масуми предложил проект многоквартирного дома, в котором балконы квартир расположены по спирали вокруг центральной оси, и ни один из них не затеняет балкон квартиры, находящейся непосредственно под ним.
Визуальный художник Сьорд Буисман использовал филлотаксис как источник вдохновения для своих работ. Примерно с 1983 года он делает изображения филлотаксиса, которые можно найти, среди прочего, в Арнеме, Гааге, Горинхеме, Гронингене, Харлеме, Леккеркерке, Оттерло и Утрехте. В качестве модели он использовал поперечное сечение растения сельдерея.
Примечание[править | править код]
- ↑ Leonardo da Vinci (1971). Taylor, Pamela (ed.). The Notebooks of Leonardo da Vinci. New American Library. p. 121.
- ↑ Бесчастнов Н. П. Изображение растительных мотивов. — М.: Гуманитарный издательский центр ВЛАДОС,, 2008. — С. 67. — 209 с. — ISBN 978-5-457-36936-8. Архивировано 17 ноября 2022 года.
- ↑ 1 2 Марио Ливио. The golden ratio : the story of phi, the world's most astonishing number. — 1st ed. — New York: Broadway Books, 2002. — С. 110. — viii, 294 pages с. — ISBN 0-7679-0815-5, 978-0-7679-0815-3, 0-7679-0816-3, 978-0-7679-0816-0.
- ↑ Базилевская Н. А., Белоконь М. П, Щербакова А. А. Краткая история ботаники. — М.: Наука, 1968. — С. 52. — 312 с. Архивировано 17 ноября 2022 года.
- ↑ Snow, M.; Snow, R. (1934). "The interpretation of Phyllotaxis". Biological Reviews. 9 (1): 132—137. doi:10.1111/j.1469-185X.1934.tb00876.x. S2CID 86184933.
- ↑ 1 2 3 4 Фролова, 1937, с. 23.
- ↑ 1 2 Завидовская Т. С. Ботаника : анатомия и морфология: курс лекций. — М., Берлин: Directmedia, 2018-05-16. — С. 120. — 213 с. — ISBN 978-5-4475-9635-4. Архивировано 17 ноября 2022 года.
- ↑ Горянинов, 1841, с. 93.
- ↑ 1 2 3 Т. И. Серебрякова, Н. С. Воронин, А. Г, Еленевский, Т. Б. Батыгина, Н. И. Шорина, Н. П. Савиных. Ботаника с основами фитоценологии: Анатомия и морфология растений. — М.: ИКЦ "Академкнига", 2007. — 543 с. — ISBN 978-5-94628-237-6.
- ↑ 1 2 3 Горянинов, 1841, с. 92.
- ↑ Костус прекрасный (по новым данным Hellenia speciosa (J.Koenig) S.R.Dutta). Коллекции растений ЦСБС СО РАН. ЦСБС СО РАН. Дата обращения: 17 ноября 2022. Архивировано 17 ноября 2022 года.
- ↑ 1 2 Листорасположение // Энциклопедический словарь Брокгауза и Ефрона : в 86 т. (82 т. и 4 доп.). — СПб., 1890—1907.
- ↑ Тахтаджян А. Л. Палеоботаника (Выпуск 6) (Труды Ботанического института Академии наук СССР, Серия 8): Проблемы изучения ископаемой флоры угленосных отложений СССР. — Л.: Наука, 1967-12-26. — Т. VI. — С. 192. — 306 с. Архивировано 17 ноября 2022 года.
- ↑ Marloth R (1932). The Flora of South Africa. Cape Town & London: Darter Bros., Wheldon & Wesley.
- ↑ Chittenden FJ (1951). Dictionary of Gardening. Oxford: Royal Horticultural Society.
- ↑ A. D. Bell. Illustrierte Morphologie der Blütenpflanzen. — Ulmer Verlag Stuttgart, 1994. — 335 с.
- ↑ Niklas, 1992, p. 513.
- ↑ Roberts, 2007, p. 101.
Литература[править | править код]
- Фролова В. М., Раменский Л. Г. Определитель растений в нецветущем состоянии для средней части СССР. — 2-е изд. — М.: Сельхозгиз, 1937. — С. 23–24. — 431 с.
- Горянинов П. Ф. Глава III. О листьях, прилистниках и пр. // Основанiя ботаники. — СПб., 1841. — С. 92–97. — 407 с.
- Niklas, Karl J. Plant Biomechanics: An Engineering Approach to Plant Form and Function. — University of Chicago Press, 1992. — 622 p. — ISBN 978-0226586304.
- Roberts, Keith. Handbook of Plant Science. — Wiley-Interscience, 2007. — Т. 1. — 1648 p. — ISBN 978-0470057230.
Ссылки[править | править код]
| Словари и энциклопедии | |
|---|---|
| В библиографических каталогах |
| Закономерности | .jpg) | |
|---|---|---|
| Процессы | ||
| Исследователи | ||
| Связанные статьи | ||