Гликогеногенез
Гликогеногене́з — метаболический путь синтеза гликогена из глюкозы, происходящий с расходованием энергии в виде ATP и UTP. Гликогеногенез происходит во всех тканях животных, однако в основном он имеет место в печени и мышцах. Синтез гликогена происходит в период пищеварения (в абсорбтивный период, т. е. 1—2 часа после приёма углеводной пищи[1][2].
Механизм
[править | править код]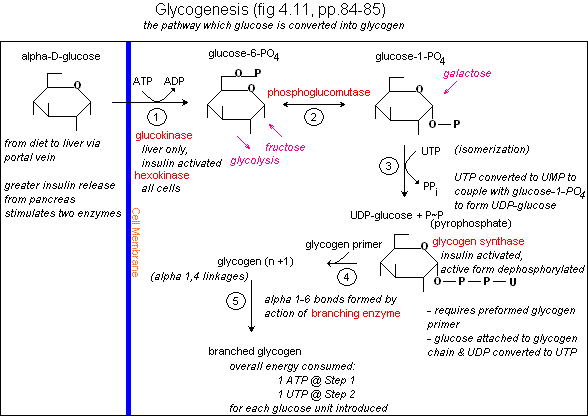{kind=link}
«Стартовой точкой» гликогеногенеза служит глюкозо-6-фосфат. Глюкозо-6-фосфат может быть получен из глюкозы в ходе реакции, катализируемой изоферментами гексокиназой I и гексокиназой II в мышцах и гексокиназой IV (глюкокиназой) в печени:
- D-глюкоза + ATP → D-глюкозо-6-фосфат + ADP.

Однако поглощённая с пищей глюкоза может проделывать более сложный путь до глюкозо-6-фосфата. Сначала она попадает в эритроциты, где гликолитически превращается в лактат. Затем лактат поступает в печень, где в ходе глюконеогенеза превращается в глюкозу, а потом и глюкозо-6-фосфат[2].
Для инициации синтеза гликогена глюкозо-6-фосфат должен быть переведён в глюкозо-1-фосфат ферментом фосфоглюкомутазой:
- Глюкозо-6-фосфат ⇌ глюкозо-1-фосфат[2].

Глюкозо-1-фосфат далее превращается в UDP-глюкозу[англ.] под действием UDP-глюкозопирофосфорилазы[англ.], это является ключевым этапом в синтезе гликогена[3].
- Глюкозо-1-фосфат + UTP → UDP-глюкоза + PPi
Эта реакция образования нуклеотид-сахара в клеточных условиях необратима, поэтому необратим и гликогеногенез. Конденсация уридинтрифосфата с глюкозо-1-фосфатом имеет небольшое положительное изменение энергии Гиббса, однако в ходе этой реакции выделяется пирофосфат (PPi), который быстро гидролизуется пирофосфатазой[англ.], а эта реакция сильно экзергонична (ΔG'o = −19,2 кДж/моль). Таким образом концентрация пирофосфата в клетке поддерживается низкой, и образование нуклеотид-сахара оказывается выгодным для клетки энергетически. В действительности быстрое вовлечение продукта реакции в другие реакции, чему способствует большое отрицательное значение изменения энергии Гиббса при гидролизе пирофосфата, стимулирует дальнейшие реакции биосинтеза[4].
UDP-глюкоза является непосредственным донором глюкозных остатков в реакции, катализируемой гликогенсинтазой[англ.], которая катализирует перенос глюкозного остатка с UDP-глюкозы на нередуцирующий конец разветвлённой молекулы гликогена[3].

Гликогенсинтаза создаёт (α1→4)-гликозидные связи, однако она неспособна создавать (α1→6)-гликозидные связи, которые располагаются в точках ветвления гликогена. Эти связи образует гликогеноразветвляющий фермент[англ.], или амило-(1→4)-(1→6)-трансгликозилаза, или гликозил-(4→6)-трансфераза. Гликогеноразветвляющий фермент катализирует перенос терминального фрагмента длиной 6 или 7 глюкозных остатков с нередуцирующего конца ветви гликогена длиной не менее 11 остатков на гидроксильную группу при шестом атоме остатка глюкозы, располагающегося ниже, причём он может принадлежать как той же самой, так и другой цепи. Таким образом создаётся новая ветвь гликогена[5].

Дополнительные остатки глюкозы могут добавляться на новую ветвь гликогена под действием гликогенсинтазы. Биологический момент разветвления молекулы гликогена состоит в том, что это повышает растворимость гликогена и увеличивает число его нередуцирующих концов, которые являются сайтами активности гликогенфосфорилазы (главный фермент гликогенолиза) и гликогенсинтазы[5].

Гликогенсинтаза не может начать синтез новой цепи гликогена с нуля. Для этого ей нужна затравка, которой может быть (α1→4)-полиглюкозная цепь или ветвь, содержащая не менее 8 глюкозных остатков. Образование затравки обеспечивает белок гликогенин[англ.], который является и местом синтеза затравки, и катализатором этого процесса. Первым этапом синтеза новой молекулы гликогена является перенос остатка глюкозы с UDP-глюкозы на гидроксильную группу остатка аминокислоты тирозина Tyr194 гликогенина, обусловленный глюкозилтрансферазной активностью белка. Растущая цепь удлиняется за счёт последовательного добавления 7 или более глюкозных остатков, каждый из которых берётся от UDP-глюкозы, эта реакция также катализируется гликогенином. На этом этапе в синтез гликогена включается гликогенсинтаза, обеспечивающая дельнейшее удлинение гликогеновой цепи. После этого гликогенин в составе β-частицы, ковалентно присоединённой к единственному нередуцирующему концу молекулы гликогена[5].
Регуляция
[править | править код]Регуляция гликогеногенеза осуществляется совместно с гликогенолизом (расщеплением гликогена) по типу переключения. Это переключение происходит при переходе из абсорбтивного состояния в постабсортивное, а также при смене состояния покоя на режим физической работы. В печени оно осуществляется при участии гормонов инсулина, глюкагона и адреналина, а в мышцах — инсулина и адреналина. Их действие на синтез и распад гликогена опосредовано изменением в противоположном направлении активности двух ключевых ферментов: гликогенсинтазы[англ.] (гликогеногенез) и гликогенфосфорилазы (гликогенолиз) при помощи их фосфорилирования/дефосфорилирования[6].
Примечания
[править | править код]- ↑ Северин, 2011, с. 241.
- ↑ 1 2 3 Nelson, Cox, 2008, p. 599.
- ↑ 1 2 Nelson, Cox, 2008, p. 600.
- ↑ Nelson, Cox, 2008, p. 598.
- ↑ 1 2 3 Nelson, Cox, 2008, p. 601.
- ↑ Северин, 2011, с. 245.
Литература
[править | править код]- David L. Nelson, Michael M. Cox. Lehninger Principles of biochemistry. — Fifth edition. — New York: W. H. Freeman and company, 2008. — 1158 p. — ISBN 978-0-7167-7108-1.
- Биологическая химия с упражнениями и задачами / Под ред. С. Е. Северина. — М.: Издательская группа «ГЭОТАР-Медиа», 2011. — 624 с.
Эта статья входит в число добротных статей русскоязычного раздела Википедии. |