Вироиды: различия между версиями
| [отпатрулированная версия] | [отпатрулированная версия] |
VAP+VYK (обсуждение | вклад) оформление |
Minina (обсуждение | вклад) Нет описания правки |
||
| Строка 14: | Строка 14: | ||
| commons = |
| commons = |
||
}} |
}} |
||
'''Виро́иды''' ({{lang-en|viroid}}) — [[Инфекция|инфекционные]] агенты, состоящие только из кольцевой [[РНК]]. Они вызывают различные [[Заболевание|заболевания]] [[Растения|растений]], в том числе {{нп5|Вироид веретеновидности клубней картофеля|веретеновидность клубней картофеля|en|Potato spindle tuber viroid}}, {{нп5|экзокортис цитрусовых||en|Citrus exocortis}} и карликовость [[Хризантема|хризантемы]]. Вироиды представляют собой ковалентно замкнутые кольцевые одноцепочечные [[молекулы]] РНК (оцРНК) длиной от 246 до 467 [[нуклеотид]]ов<ref name="Katsarou">{{cite pmid|25986218}}</ref> (для сравнения: [[геном]] мельчайшего из известных [[вирус]]ов составляет 2000 нуклеотидов в длину<ref name="Pommerville" />). В отличие от вирусов, вироиды лишены [[Белок|белкового]] [[капсид]]а. Обычно кольцевая РНК вироидов существует в виде палочковидной формы из-за [[Спаренные основания|спаривания азотистых оснований]] внутри цепи, в результате чего образуются двуцепочные участки с одноцепочечными петлями. Некоторые вироиды находятся в [[Ядрышко|ядрышке]] инфицированной [[Клетка (биология)|клетки]], где может присутствовать от 200 то 100 тысяч копий генома вироида. Другие вироиды располагаются в [[хлоропласт]]ах. РНК вироидов не кодирует каких-либо белков, поэтому вироиды не могут [[Самовоспроизведение|реплицироваться]] сами по себе. Предполагается, что для этих целей они используют фермент [[ДНК]]-зависимую [[РНК-полимераза|РНК-полимеразу]] хозяйской клетки, который обычно используется для [[синтез]]а РНК на матрице ДНК. Однако в инфицированной вироидом клетке этот [[фермент]] использует РНК вироида, а не ДНК клетки-хозяина, как матрицу для синтеза РНК. Эта молекула РНК, комплементарная геному вироида, используется как матрица для синтеза новых РНК вироида{{sfn|Willey et. al.|2011|p=105—106}}. |
|||
'''Виро́иды''' — [[патоген]]ы растений, которые состоят из короткого фрагмента (несколько сотен [[Нуклеотиды|нуклеотидов]]) [[комплементарность (биология)|высококомплементарной]], кольцевой, одноцепочечной [[Рибонуклеиновая кислота|РНК]], не покрытой белковой [[Вирусная оболочка|оболочкой]], характерной для [[Вирусы|вирусов]]. |
|||
Растение, заражённое вироидом, может не проявлять никаких [[симптом]]ов ({{нп5|Вирусная латентность|латентная инфекция|en|Virus latency}}). Однако тот же самый вироид может вызывать серьёзную болезнь у другого вида растений. Основы [[Патогенность|патогенности]] вироидов ещё не определены, однако известно, что для неё необходимы некоторые участки вироидной РНК. Некоторые данные свидетельствуют о том, что вироиды вызывают болезнь, активируя механизмы РНК-[[Сайленсинг генов|сайленсинга]] в [[Эукариоты|эукариотической]] клетке, которые в норме работают для защиты клетки от вирусов, чей геном представлен двуцепочечной РНК (дцРНК). При РНК-сайленсинге клетка распознаёт дцРНК и селективно разрушает их. Вироиды могут вмешиваться в этот процесс, [[Комплементарность|комплементарно]] связываясь ([[Гибридизация ДНК|гибридизуясь]]) со специфическими молекулами РНК клетки-хозяина. Образование гибридных дцРНК из РНК вироида и клетки запускает РНК-сайленсиг, направленный на разрушение гибридного комплекса. В результате разрушается мРНК клетки-хозяина и происходит сайленсинг её определённых [[ген]]ов. Невозможность [[Экспрессия генов|экспрессировать]] важный ген обусловливает болезнь у растения-хозяина. Впрочем, имеются предположения и о других механизмах действия вироидов на [[растительные клетки]]{{sfn|Willey et. al.|2011|p=105—106}}. |
|||
Самые малые вироиды [[scRNA]] (малые цитоплазматические РНК) вируса жёлтой крапчатости риса ({{lang-en|RYMV, rice yellow mottle sobemovirus}}) имеют длину всего 220 нуклеотидов. Для сравнения: геном самого маленького известного вируса, способного вызывать инфекцию, имеет размер около 2000 оснований. |
|||
Вироиды относятся к числу самореплицирующихся РНК-содержащих [[патоген]]ов, которые вызывают различные заболевания у растений и [[Животные|животных]]. Помимо вироидов, к таким патогенами относят сателлитные РНК, вироид-подобные растительные сателлитные РНК, а также [[Гепатит D|вирус гепатита дельта]] [[человек]]а{{sfn|Acheson|2011|p=379}}. |
|||
Вироиды были открыты и названы в [[1971 год]]у [[Динер, Теодор Отто|Теодором О. Динером]]<ref>[http://www.ars.usda.gov/is/timeline/viroid.htm Tracking the Elusive Viroid]</ref>. |
|||
== История == |
|||
[[Файл:Theodor O Diener.jpg|right|thumb|200px|Теодор Динер, первооткрыватель вироидов]] |
|||
В 1920-х годах фермеры [[Нью-Йорк (штат)|Нью-Йорка]] и [[Нью-Джерси]] заметили у [[Картофель|картофеля]] симптомы неизвестного заболевания. [[Клубень|Клубни]] поражённых растений утрачивали привычную форму и становились веретеновидными, из-за чего эта болезнь была названа веретеновидностью клубней картофеля<ref>{{cite journal | author = Owens RA, Verhoeven JT | title = Potato Spindle Tuber | journal = Plant Health Instructor | year = 2009 | doi = 10.1094/PHI-I-2009-0804-01 }}</ref>. Симптомы заболевания появлялись у растений, контактировавших с фрагментами заражённых растений, следовательно, болезнь вызывалась агентом, который мог передаваться от растения к растению. Однако у поражённых растений не обнаруживалось наличия необычного грибка или бактерии, поэтому был сделан вывод, что болезнь вызывается вирусом. Несмотря на многочисленные попытки изолировать и очистить этот вирус из [[экстракт]]а веретеновидного картофеля с использованием все более усложняющихся методов, выделить его так и не удалось<ref name = "ARS_timeline">{{cite web|url=http://www.ars.usda.gov/is/timeline/viroid.htm |title=ARS Research Timeline – Tracking the Elusive Viroid |date=2006-03-02 |accessdate=2007-07-18}}</ref>. |
|||
В 1981 году [[Динер, Теодор Отто|Теодор О. Динер]] показал, что этот инфекционный агент был не вирусом, а патогеном совершенно нового типа, размер которого составлял одну восьмидесятую от размера типичного вируса, и предложил для него термин «вироид»<ref>{{cite pmid|5095900}}</ref> (то есть «вирусоподобный»). Параллельно велись и агрокультурные исследования особенностей вироидов, и фундаментальные научные исследования, направленные на изучение физических, химических и макромолекулярных свойств вироидов. В 1976 году Сэнгер ({{lang-en|Sänger}}) и коллеги доказали, что патоген, вызывающий веретеновидность клубней картофеля, представляет собой «одноцепочечную, ковалентно замкнутую, кольцевую молекулу РНК, из-за спаривания оснований приобретающую плотную палочковидную структуру». Это было первое описание природы вироидов<ref>{{cite pmid|1069269}}</ref>. |
|||
Кольцевая форма и одноцепочечная структура молекулы РНК вироидов была подтверждена [[Электронная микроскопия|электронной микроскопией]]<ref>{{cite pmid|4728831}}</ref>, а полная последовательность генома вироида веретеновидности клубней картофеля (PSTVd) была определена в 1978 году Гроссом ({{lang-en|Gross}}) и коллегами<ref>{{cite pmid|643081}}</ref>. Этот вироид стал первым патогеном эукариот, для которого была определена полная молекулярная структура. По оценкам, более трети вирусных заболеваний растений на самом деле вызываются вироидами<ref name=Pommerville>{{cite book |last=Pommerville|first=Jeffrey C |date=2014 |title= Fundamentals of Microbiology|location=Burlington, MA |publisher= Jones and Bartlett Learning |page= 482|isbn=978-1-284-03968-9}}</ref>. |
|||
== Классификация == |
== Классификация == |
||
На основании сравнительного анализа последовательностей и наличия центрального [[Консервативные последовательности|консервативного]] участка в молекуле геномной РНК все известные на данный момент вироиды классифицируют на два [[Семейство (биология)|семейства]]. В настоящий момент известно 4 члена группы {{нп5|Вироид пятнистости авокадо|вироида пятнистости авокадо|en|Avocado sunblotch viroid}} (ASBVd), также известной как ''Avsunviroidae'' или группа А. У членов этого семейства как {{нп5|Полярность (молекулярная биология)|плюс-|en|Sense (molecular biology)}}, так и минус-цепи вироидной РНК способны самовырезаться из РНК-мультимеров. Все остальные вироиды, известные на данный момент, относятся к группе вироида веретеновидности клубней картофеля (PSTVd), также известной как ''Pospiviroidae'' или группа В. РНК этих вироидов имеет консервативный участок и неспособна к самовырезанию. Кратко основные черты семейств вироидов и охарактеризованных представителей перечислены в таблице ниже{{sfn|Acheson|2011|p=379}}. Стоит отметить, что, помимо нижеперечисленных, выделены и другие вироиды (например, латентный вироид [[Георгина|георгин]]), однако они ожидают признания [[Международный комитет по таксономии вирусов|Международным комитетом по таксономии вирусов]] (ICTV)<ref name="Katsarou" />. Если уровень гомологии выше 90 %, то сравниваемые вироиды считаются принадлежащими одному виду<ref>{{cite web| url=http://www.ictvdb.org/ICTVdB/80.htm International |title=Committee on Taxonomy of Viruses}}</ref> |
|||
Классификация вироидов основана на анализе нуклеотидной последовательности. Если уровень гомологии выше 90 %, то сравниваемые вироиды считаются принадлежащими одному виду<ref>[http://www.ictvdb.org/ICTVdB/80.htm International Committee on Taxonomy of Viruses]</ref>. Различают два семейства ''[[Pospiviroidae]]'' и ''[[Avsunviroidae]]''. Названы по сокращению первых найденных представителей: '''Po'''tato '''spi'''ndle tuber '''viroid''' (PSTVd) и '''Av'''ocado '''sun'''blotch '''viroid''' (ASBVd). ''Pospiviroidae'' отличается наличием центральной консервативной части и отсутствием в составе генома [[Рибозим типа hammerhead|рибозимов типа hammerhead]]. ''Avsunviroidae'', напротив, не имеют центральной консервативной части, но обладают в составе генома рибозимами типа hammerhead. ''Pospiviroidae'', как правило располагаются в ядре клетки генома-хозяина, а ''Avsunviroidae'' в [[хлоропласт]]е. |
|||
{|class="wide" |
|||
== Происхождение == |
|||
! Семейство !! Характерные свойства !! [[Род (биология)|Роды]] !! Представители |
|||
|- |
|||
| ''[[Avsunviroidae]]'' (группа А) || Самовырезающаяся РНК; самолигирование; репликация в хлоропластах || [[Avsunviroid]]<br/> [[Pelamoviroid]]<br/><br/>[[Elaviroid]] || Вироид пятнистости авокадо (ASBVd)<br/> {{нп5|Вироид латентной мозаичности персика||en|Peach latent mosaic viroid}} (PLMVd) и вироид хлоротичных пятен хризантемы (CChMVd)<br/> Латентный вироид баклажана (ELVd) |
|||
|- |
|||
| ''[[Pospiviroidae]]'' (группа В) || РНК неспособна к самовырезанию; репликация в [[Ядро клетки|ядре]] || {{нп5|Pospiviroid||en|Pospiviroid}}<br/> [[Cocadviroid]]<br/> {{нп5|Hostuviroid||en|Hostuviroid}}<br/> [[Apscaviroid]]<br/> [[Coleviroid]] || Вироид веретеновидности картофеля (PSTVd) и 8 других<br/> {{нп5|Вироид каданг-каданга кокосовой пальмы||en|Cadang-cadang}} (CCCVd) и 3 других<br/> {{нп5|Вироид карликовости хмеля||en|Hop stunt viroid}} (HpSVd)<br/> Вироид рубцов на кожице яблок (ASSVd) и 11 других <br/> Вироид [[Колеус Блюма|колеуса Блюма]]-1 (CbVd) и 4 других |
|||
|} |
|||
Анализ последовательностей показывает, что РНК членов семейства ''Pospiviroidae'' имеет 5 доменов: T<sub>L</sub> (терминальный левый), P (патогенный), C (центральный), V (вариабельный) и T<sub>R</sub> (терминальный правый). Границы между этими доменами определяются по резкому изменению в степени гомологии последовательностей у различных вироидов. Раньше считалось, что эти домены имеют специфичные функции, например, домен Р связан с патогенностью вироида. Однако впоследствии оказалось, что причины, лежащие в основе патогенности вироидов, более сложны, и проявление симптомов заболевания в настоящее время связывают с различными детерминантами, располагающимися в разных доменах. Члены этого семейства подразделяются на пять родов, главным образом, на основании наличия высококонсервативных последовательностей в домене С, и отчасти на основании наличия гомологий в последовательностях других доменов{{sfn|Acheson|2011|p=379—380}}. |
|||
По одной из версий считается, что вироиды представляют собой «сбежавшие [[интрон]]ы» — вырезанные в ходе [[сплайсинг]]а незначащие участки [[мРНК]], которые случайно приобрели способность к репликации. [[белки|Белков]] вироиды не [[генетический код|кодируют]]. |
|||
== Строение == |
|||
Вироиды также рассматривают в качестве представителей добиотического «[[Мир РНК|мира РНК]]», неких эволюционных реликтов<ref>Вироиды, их признаки и вредность([https://docs.google.com/document/pub?id=1EeQIAFhYW0_QMIaSyYU0g4rQj67uQsUj1vIMhz_oXsw&pli=1 html])</ref>. |
|||
{{External media |
|||
|image1 = [http://images.the-scientist.com/content/figures/0890-3670-040830-19-1-2.jpg Структура вироидов двух семейств с указанием доменов] |
|||
}} |
|||
Вироидная РНК проявляет высокую степень спаривания оснований по всей молекуле. РНК членов ''Pospiviroidae'' сворачивается в палочковидную структуру, в которой короткие двуцепочечные участки разделяются небольшими одноцепочечными петлями. Хотя предполагается, что вироид пятнистости авокадо также имеет палочковидную [[Вторичная структура|вторичную структуру]], два других члена семейства ''Avsunviroidae'', вироид латентной мозаичности [[персик]]а и вироид хлоротичных пятен хризантемы, имеют более сложные вторичные структуры, в которых от центральной (коровой) части отходят несколько [[Шпилька (биология)|шпилек]]. Реальная структура вироидов внутри клетки остаётся невыясненной, и возможно, что вироидные РНК принимают несколько альтернативных [[Конформация|конформаций]] на различных этапах жизненного цикла{{sfn|Acheson|2011|p=380}}. |
|||
[[Файл:PSTviroid.png|thumb|center|650px|Структура вироида веретеновидности клубней картофеля. Двуцепочечные участки разделяются одноцепочечными петлями]] |
|||
Принято считать, что наиболее распространённая и часто встречаемая форма вироидной РНК условно имеет положительную полярность, а комплементарная ей цепь — отрицательную. Стандартное определение полярности для РНК, при котором положительную полярность имеет цепь, кодирующая белок, в случае вироидов неприменимо, и принятые значения полярности выбраны произвольно{{sfn|Acheson|2011|p=380}}. |
|||
== Способность к репликации == |
|||
У семейства ''Avsunviroidae'' способность к репликации имеется благодаря наличию в геномной РНК кода [[Рибозим типа hammerhead|рибозима типа hammerhead]], как правило в двух вариантах с разных сторон (RZ+, RZ-). |
|||
== Жизненный цикл == |
|||
== PSTVd == |
|||
=== Общий план === |
|||
Первым идентифицированным вироидом был ''вироид клубней картофеля'' ({{lang-en|PSTVd, potato spindle tuber viroid}}). Его [[Первичная структура|первичная]] и [[Вторичная структура|вторичная структуры]]: |
|||
{{External media |
|||
|image1 = [http://jvi.asm.org/content/73/8/6353/F1.large.jpg Симметричная и асимметричная репликация вироидов] |
|||
}} |
|||
Репликация вироидов осуществляется [[Репликация по типу катящегося кольца|по механизму катящегося кольца]], при котором на кольцевой РНК, используемой в качестве матрицы, синтезируются линейные мультимеры. РНК-полимераза клетки-хозяина синтезирует на кольцевой РНК, имеющей положительную полярность, одноцепочечную линейную комплементарную минус-цепь. Поскольку на РНК-матрице нет выраженных сигналов терминации для РНК-полимеразы, транскрипция может идти по кругу в несколько циклов, в результате чего образуется линейная мультимерная РНК{{sfn|Acheson|2011|p=380}}. |
|||
У ''Avsunviroidae'' эти мультимерные РНК далее разрезаются на отдельные [[мономер]]ные минус-цепи, которые замыкаются в кольцо. Такие кольцевые минус-РНК в дальнейшем служат для аналогичного процесса — синтеза мультимерных линейных плюс-цепей РНК, которые разрезаются на мономеры, замыкающиеся в кольцо. Такой цикл может давать несколько новых дочерних плюс-цепей РНК с исходной плюс-матрицы, поскольку и в первом, и во втором случае матрица [[Транскрипция (биология)|транскрибируется]] больше, чем один раз. Такая репликация называется ''симметричной'', потому что и плюс-, и минус-цепи реплицируются одинаково{{sfn|Acheson|2011|p=380}}. |
|||
{| |
|||
Члены семейства ''Pospiviroidae'' реплицируются по схожей схеме, однако у них линейные мультимерные минус-цепи, образующиеся в первом раунде транскрипции, не разрезаются на мономеры, а непосредственно копируются в линейные мультимерные плюс-цепи. Эти плюс-мультимеры далее разрезаются на мономеры, которые замыкаются в кольцо под действием ферментов клетки, давая дочерние кольцевые молекулы положительной полярности. Такая репликация называется ''асимметричной''{{sfn|Acheson|2011|p=380}}. |
|||
Описанный выше цикл репликации по типу катящегося кольца требует работы трёх ферментов: |
|||
# {{нп5|РНК-зависимая РНК-полимераза|РНК-зависимой РНК-полимеразы|en|RNA-dependent RNA polymerase}}, которая синтезирует мультимерные линейные цепи; |
|||
# РНК-разрезающего фермента ([[Эндонуклеазы|эндорибонуклеазы]]), которая разрезает мультимерные цепи на линейные мономеры; |
|||
# РНК-[[Лигаза|лигазы]], которая замыкает линейные мономеры в кольцо{{sfn|Acheson|2011|p=380}}. |
|||
=== Полимеризация === |
|||
Считается, что в репликации ''Pospiviroidae'' участвует клеточная ДНК-зависимая [[РНК-полимераза II]] (RNAPII), поскольку репликация этих вироидов блокируется при низких концентрациях [[Альфа-аманитин|α-аманитина]], известного [[ингибитор]]а этого фермента. [[Полимеризация]] у ''Avsunviroidae'' требует участия другого фермента, так как она не останавливается под действием высоких концентраций α-аманитина. Непосредственное доказательство участие РНК-полимеразы II в репликации вироидов было получено позднее с помощью метода ко-[[Иммунопреципитация|иммунопреципитации]] вместе с доменом T<sub>L</sub> вироида, однако роль каждой из [[Белковая субъединица|субъединиц]] РНК-полимеразы II в этом процессе ещё предстоит установить<ref name="Katsarou" />. Возможно, это различие связано с различными клеточными компартментами, в которых локализуются вироиды этих двух групп: вироиды семейства ''Pospiviroidae'' находятся в ядре клетки, а ''Avsunviroidae'' — в хлоропластах, находящихся в [[Цитоплазма|цитоплазме]] клетки-хозяина. РНК-полимераза хлоропластов похожа на бактериальную РНК-полимеразу в плане устойчивости к α-аманитину, так что, возможно, в хлоропластах вироиды реплицируются, используя РНК-полимеразу хлоропластов{{sfn|Acheson|2011|p=381}}. |
|||
Клеточные РНК-полимеразы обычно транскрибируют только двуцепочечные ДНК-матрицы в РНК. Непонятно, каким образом вироиды рекрутируют эти РНК-полимеразы и заставляют их использовать свои одноцепочечные РНК-геномы в качестве матрицы для синтеза комплементарной РНК. Недавно были определены точки начала репликации и для ''Avsunviroidae'', и для ''Pospiviroidae'', так что, возможно, это поможет установить механизм рекрутирования вироидами РНК-полимераз клетки-хозяина{{sfn|Acheson|2011|p=381}}. |
|||
=== Разрезание === |
|||
{{External media |
|||
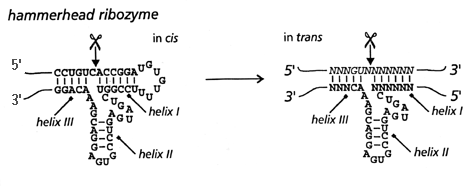
|image1 = [http://academic.brooklyn.cuny.edu/chem/zhuang/QD/2_structure.gif Структура «головка молотка»] |
|||
|image2 = [http://biowiki.ucdavis.edu/@api/deki/files/214/02hairpinribozyme1.gif?revision=1 Реакция саморазрезания] |
|||
}} |
|||
Вироиды семейства ''Avsunviroidae'' могут разрезать свои линейные мультимерные РНК на линейные мономеры ''[[in vitro]]'' в отсутствие белков. Такое разрезание молекулой РНК самой себя называется ''саморазрезанием'', а молекулы РНК, способные к саморазрезанию, называются ''[[рибозим]]ами'', так как они способны проявлять ферментативную активность в отсутствие белков{{sfn|Acheson|2011|p=381}}. |
|||
Саморазрезание у РНК ''Avsunviroidae'' осуществляется при помощи структуры «[[Рибозим типа hammerhead|головка молотка]]», которая заставляет разрыв произойти в специфической [[Фосфодиэфирная связь|фосфодиэфирной связи]], связывающей нуклеотиды РНК. Эта структура получило своё название из-за сходства с головкой молотка. Другие саморазрезающие структуры РНК были определены также у сателлитных вироид-подобных РНК (шпильки) и вируса гепатита D (дельта-структура). Все эти структуры осуществляют разрыв за счёт спаривания с основаниями в других участках молекулы РНК. Показано, что эти структуры способны образовывать мономеры из мультимеров как ''[[in vitro]]'', так и ''[[in vivo]]''. Саморазрезание происходит посредством нуклеофильной атаки, осуществляемой 2'-[[Гидроксильная группа|гидроксилом]] нуклеотида, расположенного в месте разрыва. В результате по одну сторону разрыва образуется 2',3'-циклический [[фосфат]], а по другую — свободный 5'-гидроксил{{sfn|Acheson|2011|p=381}}. |
|||
Неизвестно, как разрезаются на мономеры мультимерные плюс-цепи ''Pospiviroidae'': у них не было обнаружено способности к саморазрезанию. Было высказано предположение, что для этой цели могут использоваться клеточные ферменты{{sfn|Acheson|2011|p=382}}. Было высказано предположение, что мультимер может разрезаться на плюс-мономеры {{нп5|Рибонуклеаза III|РНКазой III|en|Ribonuclease III}}. Недавно было показано влияние [[Dicer]]-подобного белка 4 (DCL4), известного растительного белка с активностью РНКазы III, на накопление вируса веретеновидности клубней картофеля у табака ''{{нп5|Nicotiana benthamiana||en|Nicotiana benthamiana}}''<ref name="Katsarou" />. |
|||
=== Лигирование === |
|||
Лигирование у вироида латентной мозаичности персика (член семейства ''Avsunviroidae'', локализуется в хлоропластах) может происходить без участия белков клетки-хозяина. Было показано, что линейные мономеры, образующиеся при саморазрезании мультимерных молекул посредством структуры «головка молотка», способны к самолигированию ''[[in vitro]]'', образуя 3'→5'-фосфодиэфирные связи. По этой причине предполагается, что для репликации вироидам семейства ''Avsunviroidae'' необходима лишь РНК-полимераза клетки-хозяина, так как они способны к саморазрезанию и самолигированию без помощи белков{{sfn|Acheson|2011|p=382}}. |
|||
Напротив, вироидам ''Pospiviroidae'' необходимы ферменты клетки-хозяина для осуществления всех этапов жизненного цикла: полимеризации, разрезания и лигирования. Показано, что РНК-лигаза проростков пшеницы, участвующая в сплайсинге [[интрон]]ов [[тРНК]], может замыкать в кольцо линейные мономеры вироида веретеновидности клубней картофеля. Этот фермент, который образует 3'→5'-фосфодиэфирные связи, локализован в ядре, из-за чего представляется логичным кандидатом на роль фермента, осуществляющего лигирование РНК ''Pospiviroidae''{{sfn|Acheson|2011|p=382}}. Впрочем, выяснилось, что и одна РНКаза способно замыкать в кольцо мономеры этого вироида в условиях ''in vitro''. Другим возможным кандидатом на роль лигирующего фермента в жизненном цикле вироидов является ДНК-лигаза 1; возможно, в этом случае снова имеет место переключение фермента, работающего с ДНК, на РНК<ref name="Katsarou" />. |
|||
== Патогенность == |
|||
=== Пути заражения и симптомы === |
|||
{{External media |
|||
|image1 = [http://www.eppo.int/QUARANTINE/virus/PSTVd/PSTVD0_02.jpg Здоровый клубень (слева) и клубни, поражённые вироидом веретеновидности клубней картофеля] |
|||
|image2 = [http://www.dpvweb.net/dpvfigs/d402f03.jpg Кокосовая пальма, поражённая вироидом каданг-каданга] |
|||
}} |
|||
Наиболее вероятным способом передачи вироидов является передача через механические повреждения. Такой механизм передачи широко распространён среди патогенов растений. Инфекционный агент попадает в другое растение при непосредственном контакте заражённого растения с незаражённым, при использовании заражённых садовых инструментов, через [[Семя|семена]], [[Пыльца|пыльцу]] или через [[Насекомые|насекомое]]-переносчик ([[Тли|тлей]] или [[Шмели|шмелей]], однако такой способ передачи представляется сомнительным). Предполагается также возможность передачи вироида веретеновидности клубней картофеля вместе с {{нп5|Вирус скручивания листьев картофеля|вирусом скручивания листьев картофеля|en|Potato leafroll virus}} (род ''[[Polerovirus]]'' семейства {{нп5|Luteoviridae||en|Luteoviridae}}). В этом случае вироид может заключаться в вирусный капсид, так что образуется вирусная частица, содержащая внутри вироид. Это значительно облегчает распространение вироида и усложняет контроль его распространения<ref name="Katsarou" />. |
|||
Попав в новое растение, вироиды приступают к репликации и размножению, перемещаясь в другие клетки посредством межклеточных мостиков (плазмодесм). При вироидных инфекциях наблюдаются разнообразные симптомы, которые могут затрагивать как всё растение в целом, так и отдельные органы: [[лист]]ья, [[плод]]ы, [[Цветок|цветки]], [[Корень (ботаника)|корни]], органы запасания. К числу таких симптомов можно отнести обесцвечивание листьев, карликовость, появление оранжевых пятен, усиленное образование плодов, из которых лишь немногие созревают, и др{{sfn|Acheson|2011|p=382}}. На степень выраженности симптомов влияют последовательность и строение геномной РНК вироидов, состояние растения-хозяина и условия окружающей среды. Хозяевами могут быть [[Травянистые растения|травянистые]] и [[древесные растения]], овощные и [[Декоративные растения|декоративные]] культурные растения. Некоторые растения могут служить бессимптомными носителями вироидов. Например, вироид веретеновидности клубней картофеля встречается в основном в декоративных растениях семейств {{bt-ruslat|Паслёновые|Solanaceae}}, {{bt-ruslat|Норичниковые|Scrophulariaceae}} и {{bt-ruslat|Астровые|Asteraceae}}, у которых не вызывает каких-либо симптомов, однако у помидоров и картофеля он вызывает серьёзное заболевание. Возможно, что адаптация вироида к бессимптомным хозяевам вызвала малые изменения в его последовательности или структуре, что значительно увеличило степень выраженности симптомов<ref name="Katsarou" />. |
|||
=== Основы патогенности === |
|||
{{External media |
|||
|image1 = [http://www.pnas.org/content/101/9/3275/F5.large.jpg Один из предполагаемых механизмов, лежащих в основе патогенности вироидов (через РНК-интерференцию)] |
|||
}} |
|||
Несмотря на крайнюю простоту строения, вызываемые вироидами болезни растений столь же разнообразны, как вызываемые вирусами растений. Так как вироиды не кодируют белков, их действие на растение должно быть следствием неспосредственного взаимодействия вироидной РНК и содержимого клетки-хозяина. Анализ молекулярных химер, сконструированных из вироидов семейства ''Pospiviroidae'' с различной выраженностью патогенности, показал, что выраженность симптомов заболеваний зависит от сложных взаимодействий, в которых участвуют три из пяти доменов вироидной РНК. Изменения последовательности нуклеотидов в домене патогенности (Р) может изменить инфективность вироида и степень выраженности симптомов. Например, некоторые [[мутации]], усиливающие спаривание оснований в этом домене, снижают степень проявления симптомов{{sfn|Acheson|2011|p=382}}. |
|||
Молекулярный механизм, с помощью которого вироиды вызывают заболевания у растений, остаётся таинственным. Было высказано предположение, что первыми мишенями вироида являются и [[нуклеиновые кислоты]], и белки клетки-хозяина. Геномы некоторых вироидов содержат участки, комплементарные некоторым клеточным РНК. В связи с этим предполагается, что заболевание начинается из-за ингибирования функций этих клеточных РНК или их разрезания, направляемого вироидной РНК (например, ''[[Транс-регуляторные элементы|транс]]''-разрезания структурой «головка молотка»). Например, последовательность части РНК вироида веретеновидности клубней картофеля имеет сходства с последовательностью РНК U1 [[Млекопитающие|млекопитающих]] (эта РНК участвует в [[сплайсинг]]е), а некоторые вироиды могут комплементарно спариваться с 7S [[рРНК]]. Однако сложно объяснить, ка изменение нескольких нуклеотидов может превратить высокопатогенный вироид в слабый, так как эти изменения обычно не располагаются в тех участках, которые, как предполагаются, гибридизуются с клеточными РНК{{sfn|Acheson|2011|p=382}}. |
|||
Патогенность вироидов может также быть результатом [[Мимикрия|мимикрии]] на молекулярном уровне. Из-за особенностей структуры или последовательности нуклеотидов вироидная РНК может замещать собой некоторые клеточные РНК. Так, были обнаружены [[Гомология (биология)|гомологии]] между вироидом веретеновидности клубней картофеля и [[интрон]]ами I группы, а также РНК U3B, участвующими в сплайсинге РНК. Поэтому вироидные РНК могут мешать прохождению сплайсинга, замещая функциональные клеточные РНК в сплайсирующих комплексах{{sfn|Acheson|2011|p=382}}. |
|||
В вироидном патогенезе также может быть задействована [[РНК-интерференция]]. Растения используют механизмы РНК-сайленсинга для защиты от вирусных инфекций. Ферменты клетки могут распознавать чужеродные двуцепочечные РНК или одноцепочечные РНК с развитой пространственной структурой и разрезать их на [[малые интерферирующие РНК]] (siРНК) длиной 21—26 нуклеотид. В заражённых растениях были выявлены малые РНК, идентичные участкам вироидной РНК, и показано, что эти siРНК образовались в результате работы ферментов клетки после проникновения вируса. Развитие симптомов вироидной инфекции может быть обусловлено активацией или отрицательной регуляцией клеточных генов-мишеней под действием siРНК, однако к настоящему моменту не было идентифицировано специфических генов-мишеней{{sfn|Acheson|2011|p=382—383}}. |
|||
С другой стороны, возможно, существуют белки клетки-хозяина, которые распознают и взаимодействуют с различными вироидными структурами. Двуцепочечная РНК-зависимая [[протеинкиназа]] (PKR) млекопитающих активируется вироидом веретеновидности клубней картофеля, а её вторичная структура напоминает таковую двуцепочечной РНК. Была показана связь между уровнем активации этого фермента и степенью выраженности симптомов заболевания у растения. Активированная PKR [[Фосфорилирование|фосфорилирует]] альфа-субъединицу эукариотического фактора инициации синтеза белка ({{нп5|eIF2||en|eIF2}}), в результате чего происходит ингибирование синтеза белка в клетке. Активация растительного гомолога PKR может запускать вироидный [[патогенез]], поскольку в клетках млекопитающих активность PKR индуцируется [[интерферон]]ами и активируется двуцепочечной РНК{{sfn|Acheson|2011|p=382}}. |
|||
Развитие вироидной инфекции может зависеть и от других белков клетки-хозяина. Взаимодействие белков клетки с вироидами чрезвычайно сложно, потому что высокая частота мутаций вироидов может оказывать значительное действие на их геномную последовательность и/или структуру. В 2003 году был открыт [[бромодомен]]-содержащий белок [[Относительная молекулярная масса|массой]] 65 кДа (VIRP1/BRP1), имеющий {{нп5|РНК-связывающие белки|РНК-связывающий домен|en|RNA-binding protein}} и предполагаемый {{нп5|сигнал ядерной локализации||en|Nuclear localization sequence}} (NLS). Этот белок может взаимодействовать с доменом T<sub>R</sub> вироида веретеновидности корней картофеля (PSTVd) и в меньшей степени — вироида карликовости хмеля (HSVd). Показано, что у растений табака ''N. benthamiana'' и ''[[Табак обыкновенный|N. tabacum]]'', у которых была подавлена экспрессия VIRP1, от PSTVd и вироида экзокортиса цитрусовых (CEVd) инфекция не развивалась. Эти данные, а также ядерная локализация белка свидетельствуют о его возможной роли в доставке вироида в ядро. Другим интересным примером взаимодействия вироида с белками клетки-хозяина является взаимодействие с очень многочисленным [[Флоэма|флоэмным]] белком, содержащим РНК-связывающий домен — РР2, [[Лектины|лектин]] массой 49 кДа. Этот белок взаимодействует с различными молекулами РНК, в том числе HSVd (как ''in vitro'', так и ''in vivo''); предполагается, что он участвует в перемещении вироидов на дальние расстояния. Недавно открытый высокоструктрированный белок массой 30 кДа, Nt-4/1, может влиять на накопление и перемещение PSTVd. Стоит отметить, что непосредственное взаимодействие вироидов с такими белками, как [[гистоны]], {{нп5|TFIIA||en|Transcription factor II A}} и eIF1A вовлекает вироиды в регуляцию [[хроматин]]а, транскрипции и [[Трансляция (биология)|трансляции]] белков клетки-хозяина<ref name="Katsarou" />. |
|||
В таблице ниже кратко перечислены имеющиеся данные о взаимодействии вироидов с белками клетки-хозяина<ref name="Katsarou" />. |
|||
{|class="wide" |
|||
! Вироид !! Белок !! Функция белка у растения || Предполагаемая функция белка<br/> у вироида |
|||
|- |
|- |
||
| PSTVd (''Pospiviroidae'') || VIRP1/BRP1<br/> RNAPII<br/> ДНК-лигаза I<br/> eEIF1A<br/> TFIIA<br/> L5<br/> Nt-4/1<br/> Гистоны || ?<br/> Транскрипция<br/> Лигирование ДНК<br/> Трансляция<br/> Транскрипция<br/> Трансляция<br/> ?<br/> Упаковка ДНК в [[нуклеосомы]] || Ядерный импорт и/или системная инфекция<br/> Репликация<br/> Лигирование РНК<br/> ?<br/> ?<br/> ?<br/> Участие в системном перемещении<br/> ? |
|||
|align=center|'''Первичная структура''' |
|||
|- |
|- |
||
| CEVd (''Pospiviroidae'') || eEIF1A<br/> RNAPII<br/> VIRP1/BRP1 || Трансляция<br/> Транскрипция<br/> ? || ?<br/> Репликация<br/> Ядерный транспорт и/или системная инфекция |
|||
| |
|||
<br clear=all /> |
|||
1 CGGAACUAAA CUCGUGGUUC CUGUGGUUCA CACCUGACCU CCUGAGCAGA AAAGAAAAAA |
|||
61 GAAGGCGGCU CGGAGGAGCG CUUCAGGGAU CCCCGGG'''GAA AC'''CUGGAGCG AACUGGCAAA |
|||
121 AAAGGACGGU GGGGAGUGCC CAGCGGCCGA CAGGAGUAAU UCCCGCCGAA ACAGGGUUUU |
|||
181 CACCCUUCCU UUCUUCGGGU GUCCUUCCUC GCGCCCGCAG GACCACCCCU CGCCCCCUUU |
|||
241 GCGCUGUCGC UUCGGCUACU ACCCGGUGGA AACAACUGAA GCUCCCGAGA ACCGCUUUUU |
|||
301 CUCUAUCUUA CUUGCUUCGG GGCGAGGGUG UUUAGCCCUU GGAACCGCAG UUGGUUCCU |
|||
|- |
|- |
||
| HSVd (''Pospiviroidae'') || CsPP2<br/> VIRP1/BRP1 || Перемещение белков<br/> по флоэме<br/> ? || Перемещение на дальние расстояния<br/> Ядерный транспорт и/или системная инфекция |
|||
|align=center|'''Вторичная структура''' |
|||
|- |
|- |
||
| PMLVd (''Avsunviroidae'') || eEIF1A<br/> β-1,3-глюканаза<br/> {{нп5|Аминометилтрансфераза||en|Aminomethyltransferase}}<br/> Предполагаемый [[Шапероны|шаперон]]<br/> [[Динамин]]<br/> L5 || Трансляция<br/> Структура [[Клеточная стенка|клеточной стенки]]<br/> Фермент<br/> ?<br/> [[Эндоцитоз]]<br/> Трансляция || ?<br/> ?<br/> ?<br/> ?<br/> ?<br/> ? |
|||
| |
|||
<br clear=all /> |
|||
[[Файл:PSTviroid.png|thumb|center|800px|Предполагаемая вторичная структура вироида PSTVd]] |
|||
|- |
|- |
||
| ASBVd (''Avsunviroidae'') || Флоэмный лектин PP2<br/> PARBP33<br/> тРНК-лигаза<br/> CmmLec17 || Перемещение белков по флоэме<br/> Транскрипция в хлоропластах<br/> Процессинг тРНК<br/> Перемещение белков по флоэме<br/> || Перемещение на дальние расстояния<br/> Облегчение саморазрезания<br/> Замыкание в кольцо<br/> Перемещение на дальние расстояния |
|||
|} |
|} |
||
=== Кросс-защита === |
|||
== См. также == |
|||
Явление кросс-защиты имеет место между различными штаммами одного вироида или вироидами с очень близкими геномными последовательностями. Растение, инфицированное одним вироидом, не даёт возможность реплицироваться и вызывать заболевание другому вироиду. Это напоминает вирусную интерференцию, при которой присутствие в клетке одного вируса подавляет репликацию другого вируса. Механизм кросс-защиты у вироидов неизвестен. Согласно одной из гипотез, для репликации, транспорта из клетки в клетку и накопления необходим ограниченный фактор хозяйской клетки. Различное сродство вироидных РНК к этому вироиду может определять, какой из вироидов, одновременно попавших в клетку, будет преобладать; кроме того, взаимодействие вироидной РНК с этим фактором может определять его патогенность{{sfn|Acheson|2011|p=383}}. |
|||
* [[Рибозим типа hammerhead]] |
|||
* [[РНК-интерференция]] |
|||
== Некоторые вироид-подобные агенты == |
|||
* [[Прионы]] |
|||
=== Вироид-подобные сателлитные РНК растений === |
|||
* [[Вирусы]] |
|||
Растительные сателлитные РНК — это небольшие одноцепочечные РНК, репликация и заключение в капсид которых зависит от вспомогательного вируса, с которым, однако, они не имеют или практически не имеют сходства последовательностей. Более того, сателлитные РНК не необходимы для репликации вспомогательного вируса, и большинство из них не кодируют каких-либо белков. Сателлитные РНК могут иметь линейную или кольцевую форму. Подобно вироидам, в структуре сателлитных РНК велико спаривание оснований. В отличие от вироидов, они имеют капсид и потому могут более эффективно передаваться от растения к растению. Более того, подобно ''Avsunviroidae'', все вироид-подобные кольцевые растительные сателлитные РНК имеют структуры типа шпилек и головки молотка, которые катализируют разрезание мультимерных РНК на мономеры в ходе репликации по типу катящегося кольца{{sfn|Acheson|2011|p=383}}. |
|||
* [[Плазмиды]] |
|||
* [[Молекулярные информационные агенты]] |
|||
Ряд РНК-содержащих вирусов растений поддерживает репликацию сателлитных РНК, предположительно, давая собственную РНК-зависимую РНК-полимеразу для репликации сателлитных РНК и капсидные белки для формирования капсида сателлитных РНК. Присутствие сателлитных РНК может оказывать влияние на репликацию соответствующего вспомогательного вируса и модулировать симптомы вызываемого им заболевания. Судя по имеющимся сходствам, вироиды и вироид-подобные сатиллитные РНК произошли от общего предка{{sfn|Acheson|2011|p=383}}. |
|||
=== Вирус гепатита дельта === |
|||
Вирус гепатита дельта (HDV) — уникальный патоген человека, который имеет ряд общих черт с вироидами и вироид-подобными сателлитными РНК растений. Было высказано предположение, что HDV произошёл от примитивной вироид-подобной РНК через захват клеточного транскрипта. Этот патоген, передающийся с [[кровь]]ю, реплицируется в печени и часто вызывает скоротечный гепатит у [[Приматы|приматов]] и других млекопитающих. Вирус гепатита дельта также может быть связан с развитием [[Рак печени|рака печени]]. Он обнаруживается только в присутствие вируса гепатита В и использует белок оболочки [[Вирус гепатита В|вируса гепатита В]] (S-[[антиген]]) для упаковки своего генома, представленного РНК. Поскольку вирусу гепатита дельта необходим вирусный белок для упаковки, он считается РНК-сателлитом вируса гепатита В. По всему миру этим патогеном заражено более чем 15 млн людей, поэтому гепатит, вызываемый вирусом гепатита дельта, представляет собой серьёзную проблему для здравоохранения{{sfn|Acheson|2011|p=383}}. |
|||
== Вироиды и гипотеза мира РНК == |
|||
В 1989 году Т. Динер предположил, что уникальные свойства вироидов делают их более похожими на «живые ископаемые» из гипотетического доклеточного [[Мир РНК|РНК-мира]], чем интроны или другие РНК<ref>{{cite pmid|2480600}}</ref>. В таком случае вироиды, помимо вирусологии растений, имеют значение ещё и для [[Эволюционная биология|эволюционной биологии]], поскольку их свойства делают их более похожими, по сравнению с другими современными РНК, на те РНК, которые были ключевым этапом в переходе от неживой материи к живой ([[абиогенез]]). Эти свойства вироидов таковы: |
|||
# Вироиды очень малы, что облегчает репликацию; |
|||
# РНК вироидов имеет большое содержание [[гуанин]]а (G) и [[цитозин]]а (С), что увеличивает их стабильность и точность репликации; |
|||
# Кольцевая структура, что позволяет репликации проходить полностью в отсутствие поледовательностей-меток; |
|||
# Существование периодичности в структуре, что допускает их сборку в качестве модульных единиц в более крупные геномы; |
|||
# Вироиды не кодируют белки, что делает возможным их существование в среде без [[Рибосома|рибосом]]; |
|||
# У некоторых вироидов в репликации участвуют рибозимы — характерные элементы РНК-мира. |
|||
Впрочем, гипотеза Динера была забыта вплоть до 2014 года, когда Флорес и коллеги в своём обзоре опубликовали доказательства этой гипотезы (эти доказательства приведены выше)<ref>{{cite pmid|25002087}}</ref>. |
|||
== Значение == |
|||
Вызывая заболевания экономически важных культурных и декоративных растений, вироиды оказывают большое влияние на мировое сельское хозяйство. К настоящему моменту вироидные заболевания распространены на всех континентах, на которых имеют различное значение в зависимости от растения-хозяина и местных фитосанитарных мер. В 2014 году Европейская и Средиземноморская организация по защите растений ({{lang-en|European and Mediterranean plant protection organization, EPPO}}) включила в список патогенов растений, требующих объявления карантина, три вида вироидов из семейства ''Pospiviroidae'': вироид каданг-каданга кокосовой пальмы, вироид карликовости хризантемы и вироид веретеновидности клубней картофеля. Ещё один вид, вироид апикальной карликовости помидоров, попал в список патогенов, вызывающих серьёзные опасения<ref name="Katsarou" />. |
|||
== Методы == |
|||
Первым методом, который использовался для быстрой (в течение двух-трёх дней) идентификации растений, заражённых вироидами, был [[полиакриламид]]ный гель-[[электрофорез]] (PAGE). Поскольку это — единственный диагностический метод, для работы которого нет необходимости знать последовательность генома вироида, PAGE в [[Денатурация|денатурирующих]] условиях и сейчас служит основным методом идентификации новых вироидов. С начала 1980-х для рутинной идентификации стал активно применяться метод гибридизационных {{нп5|Точечный блот|точечных блотов|en|Dot blot}}, который постепенно вытеснил PAGE. Приблизительно через 10 лет для работы с вироидами стала применяться [[ПЦР с обратной транскрипцией|полимеразная цепная реакция с обратной транскрипцией]]<ref>{{cite pmid|22678585}}</ref><ref>{{статья |заглавие=A comparison of molecular methods for the routine detection of viroids. |автор =R.A. Mumford, K. Walsh & N. Boonham |ссылка=http://www.researchgate.net/profile/Rick_Mumford/publication/230175755_A_comparison_of_molecular_methods_for_the_routine_detection_of_viroids*/links/543ccf170cf20af5cfbf746c.pdf }}</ref>. |
|||
В настоящее время вироиды используются для изучения эволюционных связей между РНК- и ДНК-геномами. Они также являются идеальными биологическими молекулами для изучения связей структуры и функций молекул РНК{{sfn|Acheson|2011|p=385}}. |
|||
== Примечания == |
== Примечания == |
||
{{примечания}} |
{{примечания|2}} |
||
== |
== Литература == |
||
* {{книга |
|||
* [http://mbi.osu.edu/2005/ws2materials/elena.pdf Лекция CSIC-UPV, Испания] |
|||
| автор = Nicholas H. Acheson |
|||
| заглавие = Fundamentals of Molecular Virology |
|||
| ссылка = |
|||
| викитека = |
|||
| ответственный = |
|||
| издание = 2nd edition. |
|||
| место = |
|||
| издательство = WILEY (John Wiley & Sons, Inc.) |
|||
| год = 2011 |
|||
| pages = 379—383 |
|||
| columns = |
|||
| allpages = 528 |
|||
| серия = |
|||
| isbn = 978-0-470-90059-8 |
|||
| тираж = |
|||
| ref = Acheson |
|||
}} |
|||
* {{книга |
|||
| автор = Joanne M. Willey, Linda M. Sherwood, Christopher J. Woolverton |
|||
| заглавие = Prescott's Principles of Microbiology |
|||
| оригинал = |
|||
| ссылка = |
|||
| викитека = |
|||
| ответственный = |
|||
| издание = 1st edition |
|||
| место = |
|||
| издательство = McGraw-Hill Higher Education |
|||
| год = 2009 |
|||
| volume = |
|||
| pages = 105—106 |
|||
| columns = |
|||
| allpages = 968 |
|||
| серия = |
|||
| isbn = 978-0-07-337523-6 |
|||
| тираж = |
|||
| ref = Willey et. al. |
|||
}} |
|||
{{virusology-stub}} |
|||
{{Субвирусные частицы}} |
{{Субвирусные частицы}} |
||
{{Виды РНК}} |
{{Виды РНК}} |
||
Версия от 20:43, 3 июня 2015
| Вироиды | ||||||||
|---|---|---|---|---|---|---|---|---|
| | ||||||||
| Научная классификация | ||||||||
| Международное научное название | ||||||||
| Viroids | ||||||||
| Семейства | ||||||||
| ||||||||
Виро́иды (англ. viroid) — инфекционные агенты, состоящие только из кольцевой РНК. Они вызывают различные заболевания растений, в том числе веретеновидность клубней картофеля?!, экзокортис цитрусовых[англ.] и карликовость хризантемы. Вироиды представляют собой ковалентно замкнутые кольцевые одноцепочечные молекулы РНК (оцРНК) длиной от 246 до 467 нуклеотидов[2] (для сравнения: геном мельчайшего из известных вирусов составляет 2000 нуклеотидов в длину[3]). В отличие от вирусов, вироиды лишены белкового капсида. Обычно кольцевая РНК вироидов существует в виде палочковидной формы из-за спаривания азотистых оснований внутри цепи, в результате чего образуются двуцепочные участки с одноцепочечными петлями. Некоторые вироиды находятся в ядрышке инфицированной клетки, где может присутствовать от 200 то 100 тысяч копий генома вироида. Другие вироиды располагаются в хлоропластах. РНК вироидов не кодирует каких-либо белков, поэтому вироиды не могут реплицироваться сами по себе. Предполагается, что для этих целей они используют фермент ДНК-зависимую РНК-полимеразу хозяйской клетки, который обычно используется для синтеза РНК на матрице ДНК. Однако в инфицированной вироидом клетке этот фермент использует РНК вироида, а не ДНК клетки-хозяина, как матрицу для синтеза РНК. Эта молекула РНК, комплементарная геному вироида, используется как матрица для синтеза новых РНК вироида[4].
Растение, заражённое вироидом, может не проявлять никаких симптомов (латентная инфекция[англ.]). Однако тот же самый вироид может вызывать серьёзную болезнь у другого вида растений. Основы патогенности вироидов ещё не определены, однако известно, что для неё необходимы некоторые участки вироидной РНК. Некоторые данные свидетельствуют о том, что вироиды вызывают болезнь, активируя механизмы РНК-сайленсинга в эукариотической клетке, которые в норме работают для защиты клетки от вирусов, чей геном представлен двуцепочечной РНК (дцРНК). При РНК-сайленсинге клетка распознаёт дцРНК и селективно разрушает их. Вироиды могут вмешиваться в этот процесс, комплементарно связываясь (гибридизуясь) со специфическими молекулами РНК клетки-хозяина. Образование гибридных дцРНК из РНК вироида и клетки запускает РНК-сайленсиг, направленный на разрушение гибридного комплекса. В результате разрушается мРНК клетки-хозяина и происходит сайленсинг её определённых генов. Невозможность экспрессировать важный ген обусловливает болезнь у растения-хозяина. Впрочем, имеются предположения и о других механизмах действия вироидов на растительные клетки[4].
Вироиды относятся к числу самореплицирующихся РНК-содержащих патогенов, которые вызывают различные заболевания у растений и животных. Помимо вироидов, к таким патогенами относят сателлитные РНК, вироид-подобные растительные сателлитные РНК, а также вирус гепатита дельта человека[5].
История

В 1920-х годах фермеры Нью-Йорка и Нью-Джерси заметили у картофеля симптомы неизвестного заболевания. Клубни поражённых растений утрачивали привычную форму и становились веретеновидными, из-за чего эта болезнь была названа веретеновидностью клубней картофеля[6]. Симптомы заболевания появлялись у растений, контактировавших с фрагментами заражённых растений, следовательно, болезнь вызывалась агентом, который мог передаваться от растения к растению. Однако у поражённых растений не обнаруживалось наличия необычного грибка или бактерии, поэтому был сделан вывод, что болезнь вызывается вирусом. Несмотря на многочисленные попытки изолировать и очистить этот вирус из экстракта веретеновидного картофеля с использованием все более усложняющихся методов, выделить его так и не удалось[7].
В 1981 году Теодор О. Динер показал, что этот инфекционный агент был не вирусом, а патогеном совершенно нового типа, размер которого составлял одну восьмидесятую от размера типичного вируса, и предложил для него термин «вироид»[8] (то есть «вирусоподобный»). Параллельно велись и агрокультурные исследования особенностей вироидов, и фундаментальные научные исследования, направленные на изучение физических, химических и макромолекулярных свойств вироидов. В 1976 году Сэнгер (англ. Sänger) и коллеги доказали, что патоген, вызывающий веретеновидность клубней картофеля, представляет собой «одноцепочечную, ковалентно замкнутую, кольцевую молекулу РНК, из-за спаривания оснований приобретающую плотную палочковидную структуру». Это было первое описание природы вироидов[9].
Кольцевая форма и одноцепочечная структура молекулы РНК вироидов была подтверждена электронной микроскопией[10], а полная последовательность генома вироида веретеновидности клубней картофеля (PSTVd) была определена в 1978 году Гроссом (англ. Gross) и коллегами[11]. Этот вироид стал первым патогеном эукариот, для которого была определена полная молекулярная структура. По оценкам, более трети вирусных заболеваний растений на самом деле вызываются вироидами[3].
Классификация
На основании сравнительного анализа последовательностей и наличия центрального консервативного участка в молекуле геномной РНК все известные на данный момент вироиды классифицируют на два семейства. В настоящий момент известно 4 члена группы вироида пятнистости авокадо[англ.] (ASBVd), также известной как Avsunviroidae или группа А. У членов этого семейства как плюс-[англ.], так и минус-цепи вироидной РНК способны самовырезаться из РНК-мультимеров. Все остальные вироиды, известные на данный момент, относятся к группе вироида веретеновидности клубней картофеля (PSTVd), также известной как Pospiviroidae или группа В. РНК этих вироидов имеет консервативный участок и неспособна к самовырезанию. Кратко основные черты семейств вироидов и охарактеризованных представителей перечислены в таблице ниже[5]. Стоит отметить, что, помимо нижеперечисленных, выделены и другие вироиды (например, латентный вироид георгин), однако они ожидают признания Международным комитетом по таксономии вирусов (ICTV)[2]. Если уровень гомологии выше 90 %, то сравниваемые вироиды считаются принадлежащими одному виду[12]
| Семейство | Характерные свойства | Роды | Представители |
|---|---|---|---|
| Avsunviroidae (группа А) | Самовырезающаяся РНК; самолигирование; репликация в хлоропластах | Avsunviroid Pelamoviroid Elaviroid |
Вироид пятнистости авокадо (ASBVd) Вироид латентной мозаичности персика[англ.] (PLMVd) и вироид хлоротичных пятен хризантемы (CChMVd) Латентный вироид баклажана (ELVd) |
| Pospiviroidae (группа В) | РНК неспособна к самовырезанию; репликация в ядре | Pospiviroid[англ.] Cocadviroid Hostuviroid[англ.] Apscaviroid Coleviroid |
Вироид веретеновидности картофеля (PSTVd) и 8 других Вироид каданг-каданга кокосовой пальмы[англ.] (CCCVd) и 3 других Вироид карликовости хмеля[англ.] (HpSVd) Вироид рубцов на кожице яблок (ASSVd) и 11 других Вироид колеуса Блюма-1 (CbVd) и 4 других |
Анализ последовательностей показывает, что РНК членов семейства Pospiviroidae имеет 5 доменов: TL (терминальный левый), P (патогенный), C (центральный), V (вариабельный) и TR (терминальный правый). Границы между этими доменами определяются по резкому изменению в степени гомологии последовательностей у различных вироидов. Раньше считалось, что эти домены имеют специфичные функции, например, домен Р связан с патогенностью вироида. Однако впоследствии оказалось, что причины, лежащие в основе патогенности вироидов, более сложны, и проявление симптомов заболевания в настоящее время связывают с различными детерминантами, располагающимися в разных доменах. Члены этого семейства подразделяются на пять родов, главным образом, на основании наличия высококонсервативных последовательностей в домене С, и отчасти на основании наличия гомологий в последовательностях других доменов[13].
Строение
{kind=link}
Вироидная РНК проявляет высокую степень спаривания оснований по всей молекуле. РНК членов Pospiviroidae сворачивается в палочковидную структуру, в которой короткие двуцепочечные участки разделяются небольшими одноцепочечными петлями. Хотя предполагается, что вироид пятнистости авокадо также имеет палочковидную вторичную структуру, два других члена семейства Avsunviroidae, вироид латентной мозаичности персика и вироид хлоротичных пятен хризантемы, имеют более сложные вторичные структуры, в которых от центральной (коровой) части отходят несколько шпилек. Реальная структура вироидов внутри клетки остаётся невыясненной, и возможно, что вироидные РНК принимают несколько альтернативных конформаций на различных этапах жизненного цикла[14].

Принято считать, что наиболее распространённая и часто встречаемая форма вироидной РНК условно имеет положительную полярность, а комплементарная ей цепь — отрицательную. Стандартное определение полярности для РНК, при котором положительную полярность имеет цепь, кодирующая белок, в случае вироидов неприменимо, и принятые значения полярности выбраны произвольно[14].
Жизненный цикл
Общий план
{kind=link}
Репликация вироидов осуществляется по механизму катящегося кольца, при котором на кольцевой РНК, используемой в качестве матрицы, синтезируются линейные мультимеры. РНК-полимераза клетки-хозяина синтезирует на кольцевой РНК, имеющей положительную полярность, одноцепочечную линейную комплементарную минус-цепь. Поскольку на РНК-матрице нет выраженных сигналов терминации для РНК-полимеразы, транскрипция может идти по кругу в несколько циклов, в результате чего образуется линейная мультимерная РНК[14].
У Avsunviroidae эти мультимерные РНК далее разрезаются на отдельные мономерные минус-цепи, которые замыкаются в кольцо. Такие кольцевые минус-РНК в дальнейшем служат для аналогичного процесса — синтеза мультимерных линейных плюс-цепей РНК, которые разрезаются на мономеры, замыкающиеся в кольцо. Такой цикл может давать несколько новых дочерних плюс-цепей РНК с исходной плюс-матрицы, поскольку и в первом, и во втором случае матрица транскрибируется больше, чем один раз. Такая репликация называется симметричной, потому что и плюс-, и минус-цепи реплицируются одинаково[14].
Члены семейства Pospiviroidae реплицируются по схожей схеме, однако у них линейные мультимерные минус-цепи, образующиеся в первом раунде транскрипции, не разрезаются на мономеры, а непосредственно копируются в линейные мультимерные плюс-цепи. Эти плюс-мультимеры далее разрезаются на мономеры, которые замыкаются в кольцо под действием ферментов клетки, давая дочерние кольцевые молекулы положительной полярности. Такая репликация называется асимметричной[14].
Описанный выше цикл репликации по типу катящегося кольца требует работы трёх ферментов:
- РНК-зависимой РНК-полимеразы?!, которая синтезирует мультимерные линейные цепи;
- РНК-разрезающего фермента (эндорибонуклеазы), которая разрезает мультимерные цепи на линейные мономеры;
- РНК-лигазы, которая замыкает линейные мономеры в кольцо[14].
Полимеризация
Считается, что в репликации Pospiviroidae участвует клеточная ДНК-зависимая РНК-полимераза II (RNAPII), поскольку репликация этих вироидов блокируется при низких концентрациях α-аманитина, известного ингибитора этого фермента. Полимеризация у Avsunviroidae требует участия другого фермента, так как она не останавливается под действием высоких концентраций α-аманитина. Непосредственное доказательство участие РНК-полимеразы II в репликации вироидов было получено позднее с помощью метода ко-иммунопреципитации вместе с доменом TL вироида, однако роль каждой из субъединиц РНК-полимеразы II в этом процессе ещё предстоит установить[2]. Возможно, это различие связано с различными клеточными компартментами, в которых локализуются вироиды этих двух групп: вироиды семейства Pospiviroidae находятся в ядре клетки, а Avsunviroidae — в хлоропластах, находящихся в цитоплазме клетки-хозяина. РНК-полимераза хлоропластов похожа на бактериальную РНК-полимеразу в плане устойчивости к α-аманитину, так что, возможно, в хлоропластах вироиды реплицируются, используя РНК-полимеразу хлоропластов[15].
Клеточные РНК-полимеразы обычно транскрибируют только двуцепочечные ДНК-матрицы в РНК. Непонятно, каким образом вироиды рекрутируют эти РНК-полимеразы и заставляют их использовать свои одноцепочечные РНК-геномы в качестве матрицы для синтеза комплементарной РНК. Недавно были определены точки начала репликации и для Avsunviroidae, и для Pospiviroidae, так что, возможно, это поможет установить механизм рекрутирования вироидами РНК-полимераз клетки-хозяина[15].
Разрезание
{kind=link}
{kind=link}
Вироиды семейства Avsunviroidae могут разрезать свои линейные мультимерные РНК на линейные мономеры in vitro в отсутствие белков. Такое разрезание молекулой РНК самой себя называется саморазрезанием, а молекулы РНК, способные к саморазрезанию, называются рибозимами, так как они способны проявлять ферментативную активность в отсутствие белков[15].
Саморазрезание у РНК Avsunviroidae осуществляется при помощи структуры «головка молотка», которая заставляет разрыв произойти в специфической фосфодиэфирной связи, связывающей нуклеотиды РНК. Эта структура получило своё название из-за сходства с головкой молотка. Другие саморазрезающие структуры РНК были определены также у сателлитных вироид-подобных РНК (шпильки) и вируса гепатита D (дельта-структура). Все эти структуры осуществляют разрыв за счёт спаривания с основаниями в других участках молекулы РНК. Показано, что эти структуры способны образовывать мономеры из мультимеров как in vitro, так и in vivo. Саморазрезание происходит посредством нуклеофильной атаки, осуществляемой 2'-гидроксилом нуклеотида, расположенного в месте разрыва. В результате по одну сторону разрыва образуется 2',3'-циклический фосфат, а по другую — свободный 5'-гидроксил[15].
Неизвестно, как разрезаются на мономеры мультимерные плюс-цепи Pospiviroidae: у них не было обнаружено способности к саморазрезанию. Было высказано предположение, что для этой цели могут использоваться клеточные ферменты[16]. Было высказано предположение, что мультимер может разрезаться на плюс-мономеры РНКазой III?!. Недавно было показано влияние Dicer-подобного белка 4 (DCL4), известного растительного белка с активностью РНКазы III, на накопление вируса веретеновидности клубней картофеля у табака Nicotiana benthamiana[англ.][2].
Лигирование
Лигирование у вироида латентной мозаичности персика (член семейства Avsunviroidae, локализуется в хлоропластах) может происходить без участия белков клетки-хозяина. Было показано, что линейные мономеры, образующиеся при саморазрезании мультимерных молекул посредством структуры «головка молотка», способны к самолигированию in vitro, образуя 3'→5'-фосфодиэфирные связи. По этой причине предполагается, что для репликации вироидам семейства Avsunviroidae необходима лишь РНК-полимераза клетки-хозяина, так как они способны к саморазрезанию и самолигированию без помощи белков[16].
Напротив, вироидам Pospiviroidae необходимы ферменты клетки-хозяина для осуществления всех этапов жизненного цикла: полимеризации, разрезания и лигирования. Показано, что РНК-лигаза проростков пшеницы, участвующая в сплайсинге интронов тРНК, может замыкать в кольцо линейные мономеры вироида веретеновидности клубней картофеля. Этот фермент, который образует 3'→5'-фосфодиэфирные связи, локализован в ядре, из-за чего представляется логичным кандидатом на роль фермента, осуществляющего лигирование РНК Pospiviroidae[16]. Впрочем, выяснилось, что и одна РНКаза способно замыкать в кольцо мономеры этого вироида в условиях in vitro. Другим возможным кандидатом на роль лигирующего фермента в жизненном цикле вироидов является ДНК-лигаза 1; возможно, в этом случае снова имеет место переключение фермента, работающего с ДНК, на РНК[2].
Патогенность
Пути заражения и симптомы
{kind=link}
{kind=link}
Наиболее вероятным способом передачи вироидов является передача через механические повреждения. Такой механизм передачи широко распространён среди патогенов растений. Инфекционный агент попадает в другое растение при непосредственном контакте заражённого растения с незаражённым, при использовании заражённых садовых инструментов, через семена, пыльцу или через насекомое-переносчик (тлей или шмелей, однако такой способ передачи представляется сомнительным). Предполагается также возможность передачи вироида веретеновидности клубней картофеля вместе с вирусом скручивания листьев картофеля[англ.] (род Polerovirus семейства Luteoviridae[англ.]). В этом случае вироид может заключаться в вирусный капсид, так что образуется вирусная частица, содержащая внутри вироид. Это значительно облегчает распространение вироида и усложняет контроль его распространения[2].
Попав в новое растение, вироиды приступают к репликации и размножению, перемещаясь в другие клетки посредством межклеточных мостиков (плазмодесм). При вироидных инфекциях наблюдаются разнообразные симптомы, которые могут затрагивать как всё растение в целом, так и отдельные органы: листья, плоды, цветки, корни, органы запасания. К числу таких симптомов можно отнести обесцвечивание листьев, карликовость, появление оранжевых пятен, усиленное образование плодов, из которых лишь немногие созревают, и др[16]. На степень выраженности симптомов влияют последовательность и строение геномной РНК вироидов, состояние растения-хозяина и условия окружающей среды. Хозяевами могут быть травянистые и древесные растения, овощные и декоративные культурные растения. Некоторые растения могут служить бессимптомными носителями вироидов. Например, вироид веретеновидности клубней картофеля встречается в основном в декоративных растениях семейств Паслёновые (Solanaceae), Норичниковые (Scrophulariaceae) и Астровые (Asteraceae), у которых не вызывает каких-либо симптомов, однако у помидоров и картофеля он вызывает серьёзное заболевание. Возможно, что адаптация вироида к бессимптомным хозяевам вызвала малые изменения в его последовательности или структуре, что значительно увеличило степень выраженности симптомов[2].
Основы патогенности
{kind=link}
Несмотря на крайнюю простоту строения, вызываемые вироидами болезни растений столь же разнообразны, как вызываемые вирусами растений. Так как вироиды не кодируют белков, их действие на растение должно быть следствием неспосредственного взаимодействия вироидной РНК и содержимого клетки-хозяина. Анализ молекулярных химер, сконструированных из вироидов семейства Pospiviroidae с различной выраженностью патогенности, показал, что выраженность симптомов заболеваний зависит от сложных взаимодействий, в которых участвуют три из пяти доменов вироидной РНК. Изменения последовательности нуклеотидов в домене патогенности (Р) может изменить инфективность вироида и степень выраженности симптомов. Например, некоторые мутации, усиливающие спаривание оснований в этом домене, снижают степень проявления симптомов[16].
Молекулярный механизм, с помощью которого вироиды вызывают заболевания у растений, остаётся таинственным. Было высказано предположение, что первыми мишенями вироида являются и нуклеиновые кислоты, и белки клетки-хозяина. Геномы некоторых вироидов содержат участки, комплементарные некоторым клеточным РНК. В связи с этим предполагается, что заболевание начинается из-за ингибирования функций этих клеточных РНК или их разрезания, направляемого вироидной РНК (например, транс-разрезания структурой «головка молотка»). Например, последовательность части РНК вироида веретеновидности клубней картофеля имеет сходства с последовательностью РНК U1 млекопитающих (эта РНК участвует в сплайсинге), а некоторые вироиды могут комплементарно спариваться с 7S рРНК. Однако сложно объяснить, ка изменение нескольких нуклеотидов может превратить высокопатогенный вироид в слабый, так как эти изменения обычно не располагаются в тех участках, которые, как предполагаются, гибридизуются с клеточными РНК[16].
Патогенность вироидов может также быть результатом мимикрии на молекулярном уровне. Из-за особенностей структуры или последовательности нуклеотидов вироидная РНК может замещать собой некоторые клеточные РНК. Так, были обнаружены гомологии между вироидом веретеновидности клубней картофеля и интронами I группы, а также РНК U3B, участвующими в сплайсинге РНК. Поэтому вироидные РНК могут мешать прохождению сплайсинга, замещая функциональные клеточные РНК в сплайсирующих комплексах[16].
В вироидном патогенезе также может быть задействована РНК-интерференция. Растения используют механизмы РНК-сайленсинга для защиты от вирусных инфекций. Ферменты клетки могут распознавать чужеродные двуцепочечные РНК или одноцепочечные РНК с развитой пространственной структурой и разрезать их на малые интерферирующие РНК (siРНК) длиной 21—26 нуклеотид. В заражённых растениях были выявлены малые РНК, идентичные участкам вироидной РНК, и показано, что эти siРНК образовались в результате работы ферментов клетки после проникновения вируса. Развитие симптомов вироидной инфекции может быть обусловлено активацией или отрицательной регуляцией клеточных генов-мишеней под действием siРНК, однако к настоящему моменту не было идентифицировано специфических генов-мишеней[17].
С другой стороны, возможно, существуют белки клетки-хозяина, которые распознают и взаимодействуют с различными вироидными структурами. Двуцепочечная РНК-зависимая протеинкиназа (PKR) млекопитающих активируется вироидом веретеновидности клубней картофеля, а её вторичная структура напоминает таковую двуцепочечной РНК. Была показана связь между уровнем активации этого фермента и степенью выраженности симптомов заболевания у растения. Активированная PKR фосфорилирует альфа-субъединицу эукариотического фактора инициации синтеза белка (eIF2[англ.]), в результате чего происходит ингибирование синтеза белка в клетке. Активация растительного гомолога PKR может запускать вироидный патогенез, поскольку в клетках млекопитающих активность PKR индуцируется интерферонами и активируется двуцепочечной РНК[16].
Развитие вироидной инфекции может зависеть и от других белков клетки-хозяина. Взаимодействие белков клетки с вироидами чрезвычайно сложно, потому что высокая частота мутаций вироидов может оказывать значительное действие на их геномную последовательность и/или структуру. В 2003 году был открыт бромодомен-содержащий белок массой 65 кДа (VIRP1/BRP1), имеющий РНК-связывающий домен[англ.] и предполагаемый сигнал ядерной локализации?! (NLS). Этот белок может взаимодействовать с доменом TR вироида веретеновидности корней картофеля (PSTVd) и в меньшей степени — вироида карликовости хмеля (HSVd). Показано, что у растений табака N. benthamiana и N. tabacum, у которых была подавлена экспрессия VIRP1, от PSTVd и вироида экзокортиса цитрусовых (CEVd) инфекция не развивалась. Эти данные, а также ядерная локализация белка свидетельствуют о его возможной роли в доставке вироида в ядро. Другим интересным примером взаимодействия вироида с белками клетки-хозяина является взаимодействие с очень многочисленным флоэмным белком, содержащим РНК-связывающий домен — РР2, лектин массой 49 кДа. Этот белок взаимодействует с различными молекулами РНК, в том числе HSVd (как in vitro, так и in vivo); предполагается, что он участвует в перемещении вироидов на дальние расстояния. Недавно открытый высокоструктрированный белок массой 30 кДа, Nt-4/1, может влиять на накопление и перемещение PSTVd. Стоит отметить, что непосредственное взаимодействие вироидов с такими белками, как гистоны, TFIIA[англ.] и eIF1A вовлекает вироиды в регуляцию хроматина, транскрипции и трансляции белков клетки-хозяина[2].
В таблице ниже кратко перечислены имеющиеся данные о взаимодействии вироидов с белками клетки-хозяина[2].
| Вироид | Белок | Функция белка у растения | Предполагаемая функция белка у вироида |
|---|---|---|---|
| PSTVd (Pospiviroidae) | VIRP1/BRP1 RNAPII ДНК-лигаза I eEIF1A TFIIA L5 Nt-4/1 Гистоны |
? Транскрипция Лигирование ДНК Трансляция Транскрипция Трансляция ? Упаковка ДНК в нуклеосомы |
Ядерный импорт и/или системная инфекция Репликация Лигирование РНК ? ? ? Участие в системном перемещении ? |
| CEVd (Pospiviroidae) | eEIF1A RNAPII VIRP1/BRP1 |
Трансляция Транскрипция ? |
? Репликация Ядерный транспорт и/или системная инфекция |
| HSVd (Pospiviroidae) | CsPP2 VIRP1/BRP1 |
Перемещение белков по флоэме ? |
Перемещение на дальние расстояния Ядерный транспорт и/или системная инфекция |
| PMLVd (Avsunviroidae) | eEIF1A β-1,3-глюканаза Аминометилтрансфераза[англ.] Предполагаемый шаперон Динамин L5 |
Трансляция Структура клеточной стенки Фермент ? Эндоцитоз Трансляция |
? ? ? ? ? ? |
| ASBVd (Avsunviroidae) | Флоэмный лектин PP2 PARBP33 тРНК-лигаза CmmLec17 |
Перемещение белков по флоэме Транскрипция в хлоропластах Процессинг тРНК Перемещение белков по флоэме |
Перемещение на дальние расстояния Облегчение саморазрезания Замыкание в кольцо Перемещение на дальние расстояния |
Кросс-защита
Явление кросс-защиты имеет место между различными штаммами одного вироида или вироидами с очень близкими геномными последовательностями. Растение, инфицированное одним вироидом, не даёт возможность реплицироваться и вызывать заболевание другому вироиду. Это напоминает вирусную интерференцию, при которой присутствие в клетке одного вируса подавляет репликацию другого вируса. Механизм кросс-защиты у вироидов неизвестен. Согласно одной из гипотез, для репликации, транспорта из клетки в клетку и накопления необходим ограниченный фактор хозяйской клетки. Различное сродство вироидных РНК к этому вироиду может определять, какой из вироидов, одновременно попавших в клетку, будет преобладать; кроме того, взаимодействие вироидной РНК с этим фактором может определять его патогенность[18].
Некоторые вироид-подобные агенты
Вироид-подобные сателлитные РНК растений
Растительные сателлитные РНК — это небольшие одноцепочечные РНК, репликация и заключение в капсид которых зависит от вспомогательного вируса, с которым, однако, они не имеют или практически не имеют сходства последовательностей. Более того, сателлитные РНК не необходимы для репликации вспомогательного вируса, и большинство из них не кодируют каких-либо белков. Сателлитные РНК могут иметь линейную или кольцевую форму. Подобно вироидам, в структуре сателлитных РНК велико спаривание оснований. В отличие от вироидов, они имеют капсид и потому могут более эффективно передаваться от растения к растению. Более того, подобно Avsunviroidae, все вироид-подобные кольцевые растительные сателлитные РНК имеют структуры типа шпилек и головки молотка, которые катализируют разрезание мультимерных РНК на мономеры в ходе репликации по типу катящегося кольца[18].
Ряд РНК-содержащих вирусов растений поддерживает репликацию сателлитных РНК, предположительно, давая собственную РНК-зависимую РНК-полимеразу для репликации сателлитных РНК и капсидные белки для формирования капсида сателлитных РНК. Присутствие сателлитных РНК может оказывать влияние на репликацию соответствующего вспомогательного вируса и модулировать симптомы вызываемого им заболевания. Судя по имеющимся сходствам, вироиды и вироид-подобные сатиллитные РНК произошли от общего предка[18].
Вирус гепатита дельта
Вирус гепатита дельта (HDV) — уникальный патоген человека, который имеет ряд общих черт с вироидами и вироид-подобными сателлитными РНК растений. Было высказано предположение, что HDV произошёл от примитивной вироид-подобной РНК через захват клеточного транскрипта. Этот патоген, передающийся с кровью, реплицируется в печени и часто вызывает скоротечный гепатит у приматов и других млекопитающих. Вирус гепатита дельта также может быть связан с развитием рака печени. Он обнаруживается только в присутствие вируса гепатита В и использует белок оболочки вируса гепатита В (S-антиген) для упаковки своего генома, представленного РНК. Поскольку вирусу гепатита дельта необходим вирусный белок для упаковки, он считается РНК-сателлитом вируса гепатита В. По всему миру этим патогеном заражено более чем 15 млн людей, поэтому гепатит, вызываемый вирусом гепатита дельта, представляет собой серьёзную проблему для здравоохранения[18].
Вироиды и гипотеза мира РНК
В 1989 году Т. Динер предположил, что уникальные свойства вироидов делают их более похожими на «живые ископаемые» из гипотетического доклеточного РНК-мира, чем интроны или другие РНК[19]. В таком случае вироиды, помимо вирусологии растений, имеют значение ещё и для эволюционной биологии, поскольку их свойства делают их более похожими, по сравнению с другими современными РНК, на те РНК, которые были ключевым этапом в переходе от неживой материи к живой (абиогенез). Эти свойства вироидов таковы:
- Вироиды очень малы, что облегчает репликацию;
- РНК вироидов имеет большое содержание гуанина (G) и цитозина (С), что увеличивает их стабильность и точность репликации;
- Кольцевая структура, что позволяет репликации проходить полностью в отсутствие поледовательностей-меток;
- Существование периодичности в структуре, что допускает их сборку в качестве модульных единиц в более крупные геномы;
- Вироиды не кодируют белки, что делает возможным их существование в среде без рибосом;
- У некоторых вироидов в репликации участвуют рибозимы — характерные элементы РНК-мира.
Впрочем, гипотеза Динера была забыта вплоть до 2014 года, когда Флорес и коллеги в своём обзоре опубликовали доказательства этой гипотезы (эти доказательства приведены выше)[20].
Значение
Вызывая заболевания экономически важных культурных и декоративных растений, вироиды оказывают большое влияние на мировое сельское хозяйство. К настоящему моменту вироидные заболевания распространены на всех континентах, на которых имеют различное значение в зависимости от растения-хозяина и местных фитосанитарных мер. В 2014 году Европейская и Средиземноморская организация по защите растений (англ. European and Mediterranean plant protection organization, EPPO) включила в список патогенов растений, требующих объявления карантина, три вида вироидов из семейства Pospiviroidae: вироид каданг-каданга кокосовой пальмы, вироид карликовости хризантемы и вироид веретеновидности клубней картофеля. Ещё один вид, вироид апикальной карликовости помидоров, попал в список патогенов, вызывающих серьёзные опасения[2].
Методы
Первым методом, который использовался для быстрой (в течение двух-трёх дней) идентификации растений, заражённых вироидами, был полиакриламидный гель-электрофорез (PAGE). Поскольку это — единственный диагностический метод, для работы которого нет необходимости знать последовательность генома вироида, PAGE в денатурирующих условиях и сейчас служит основным методом идентификации новых вироидов. С начала 1980-х для рутинной идентификации стал активно применяться метод гибридизационных точечных блотов[англ.], который постепенно вытеснил PAGE. Приблизительно через 10 лет для работы с вироидами стала применяться полимеразная цепная реакция с обратной транскрипцией[21][22].
В настоящее время вироиды используются для изучения эволюционных связей между РНК- и ДНК-геномами. Они также являются идеальными биологическими молекулами для изучения связей структуры и функций молекул РНК[23].
Примечания
- ↑ Таксономия вирусов (англ.) на сайте Международного комитета по таксономии вирусов (ICTV).
- ↑ 1 2 3 4 5 6 7 8 9 10 Katsarou K., Rao A. L., Tsagris M., Kalantidis K. Infectious long non-coding RNAs. (англ.) // Biochimie. — 2015. — doi:10.1016/j.biochi.2015.05.005. — PMID 25986218.
- ↑ 1 2 Pommerville, Jeffrey C. Fundamentals of Microbiology. — Burlington, MA : Jones and Bartlett Learning, 2014. — P. 482. — ISBN 978-1-284-03968-9.
- ↑ 1 2 Willey et. al., 2011, p. 105—106.
- ↑ 1 2 Acheson, 2011, p. 379.
- ↑ Owens RA, Verhoeven JT (2009). "Potato Spindle Tuber". Plant Health Instructor. doi:10.1094/PHI-I-2009-0804-01.
- ↑ ARS Research Timeline – Tracking the Elusive Viroid (2 марта 2006). Дата обращения: 18 июля 2007.
- ↑ Diener T. O. Potato spindle tuber "virus". IV. A replicating, low molecular weight RNA. (англ.) // Virology. — 1971. — Vol. 45, no. 2. — P. 411—428. — PMID 5095900.
- ↑ Sanger H. L., Klotz G., Riesner D., Gross H. J., Kleinschmidt A. K. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 1976. — Vol. 73, no. 11. — P. 3852—3856. — PMID 1069269.
- ↑ Sogo J. M., Koller T., Diener T. O. Potato spindle tuber viroid. X. Visualization and size determination by electron microscopy. (англ.) // Virology. — 1973. — Vol. 55, no. 1. — P. 70—80. — PMID 4728831.
- ↑ Gross H. J., Domdey H., Lossow C., Jank P., Raba M., Alberty H., Sänger H. L. Nucleotide sequence and secondary structure of potato spindle tuber viroid. (англ.) // Nature. — 1978. — Vol. 273, no. 5659. — P. 203—208. — PMID 643081.
- ↑ International Committee on Taxonomy of Viruses.
- ↑ Acheson, 2011, p. 379—380.
- ↑ 1 2 3 4 5 6 Acheson, 2011, p. 380.
- ↑ 1 2 3 4 Acheson, 2011, p. 381.
- ↑ 1 2 3 4 5 6 7 8 Acheson, 2011, p. 382.
- ↑ Acheson, 2011, p. 382—383.
- ↑ 1 2 3 4 Acheson, 2011, p. 383.
- ↑ Diener T. O. Circular RNAs: relics of precellular evolution? (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 1989. — Vol. 86, no. 23. — P. 9370—9374. — PMID 2480600.
- ↑ Flores R., Gago-Zachert S., Serra P., Sanjuán R., Elena S. F. Viroids: survivors from the RNA world? (англ.) // Annual review of microbiology. — 2014. — Vol. 68. — P. 395—414. — doi:10.1146/annurev-micro-091313-103416. — PMID 25002087.
- ↑ Owens R. A., Sano T., Duran-Vila N. Plant viroids: isolation, characterization/detection, and analysis. (англ.) // Methods in molecular biology (Clifton, N.J.). — 2012. — Vol. 894. — P. 253—271. — doi:10.1007/978-1-61779-882-5_17. — PMID 22678585.
- ↑ R.A. Mumford, K. Walsh & N. Boonham. A comparison of molecular methods for the routine detection of viroids..
- ↑ Acheson, 2011, p. 385.
Литература
- Nicholas H. Acheson. Fundamentals of Molecular Virology. — 2nd edition.. — WILEY (John Wiley & Sons, Inc.), 2011. — P. 379—383. — 528 p. — ISBN 978-0-470-90059-8.
- Joanne M. Willey, Linda M. Sherwood, Christopher J. Woolverton. Prescott's Principles of Microbiology. — 1st edition. — McGraw-Hill Higher Education, 2009. — P. 105—106. — 968 p. — ISBN 978-0-07-337523-6.
Субвирусные частицы | |
|---|---|
Виды РНК | |
|---|---|
| Биосинтез белка | |
| Процессинг РНК | |
| Регуляция экспрессии генов | |
| Цис-регуляторные элементы | |
| Паразитические элементы | |
| Прочее | |