Митоз

Мито́з (др.-греч. μίτος — «нить»), или кариокине́з (от др.-греч. κάρυον — «орех» + др.-греч. κίνησις — «движение»)[1][2] — непрямое деление клетки, наиболее распространённый способ размножения эукариотических клеток. Биологическое значение митоза состоит в строго одинаковом распределении хромосом между дочерними ядрами, что обеспечивает образование генетически одинаковых дочерних клеток и сохраняет преемственность в ряду клеточных поколений[3]. Перед делением число хромосом в клетке увеличивается в два раза продольным разделением на две части каждой из них, поэтому в каждую из дочерних клеток переходит столько же хромосом, сколько их было в родительской клетке[4].
Митоз — один из фундаментальных процессов онтогенеза (жизни индивидуального организма). Митотическое деление обеспечивает рост многоклеточных эукариот за счёт увеличения популяций клеток тканей. У растений в результате митотического деления клеток образовательных тканей (меристем) увеличивается количество клеток тканей. Дробление оплодотворённого яйца и рост большинства тканей у животных также происходит путём митотических делений[5].
На основании морфологических особенностей митоз условно подразделяется на стадии: профазу, прометафазу, метафазу, анафазу, телофазу.
Продолжительность митоза в среднем составляет 1−2 часа[3][6]. Митоз клеток животных, как правило, длится 30−60 минут, а растений — 2−3 часа[7]. За 70 лет в теле человека суммарно осуществляется порядка 1014 клеточных делений[8].
Митоз происходит только в клетках эукариот (ядерных). Клетки прокариот (безъядерных) делятся другим, бинарным, способом. Митоз отличается для разных организмов[9]. Так, например, процесс для клеток животных является «открытым», а для клеток грибов — «закрытым» (при котором хромосомы делятся в целом клеточном ядре)[10]. У диплонтов, включая человека, имеющих в жизненном цикле короткую гаплоидную стадию из гамет, все клетки, кроме гамет, производятся митозом, а гаметы производятся мейозом; у гаплонтов (например, у малярийного плазмодия), у которых жизненный цикл состоит в основном из гаплоидной стадии, а диплоидной является только зигота, все клетки, включая половые, делятся митозом, а зигота делится мейозом[11].
История исследования[править | править код]
Первые описания фаз митоза и установление их последовательности были предприняты в 1870—1890-х годах. В конце 1870-х — начале 1880-х годов немецкий гистолог Вальтер Флемминг для обозначения процесса непрямого деления клетки ввёл термин «митоз»[12].
Первые неполные описания, касающиеся поведения и изменения ядер в делящихся клетках, встречаются в работах учёных начала 1870-х годов. В работе русского ботаника Эдмунда Руссова, датируемой 1872 годом, отчётливо описаны и изображены метафазные и анафазные пластинки, состоящие из отдельных хромосом[13]. Годом позже немецкий зоолог Антон Шнейдер ещё более отчётливо и последовательно, но, конечно, не совсем полно описал митотическое деление на примере дробящихся яиц Mesostoma ehrenbergii[14]. В его работе, в сущности, описаны и проиллюстрированы в правильной последовательности основные фазы митоза: профаза, метафаза, анафаза (ранняя и поздняя). В 1874 году московский ботаник И. Д. Чистяков также наблюдал отдельные фазы клеточного деления в спорах плаунов и хвощей. Несмотря на первые успехи ни Руссову, ни Шнейдеру, ни Чистякову не удалось дать чёткое и последовательное описание митотического деления[15].
В 1875 году вышли работы, содержащие более детальные описания митозов. Отто Бючли дал описание цитологических картин в дробящихся яйцах круглых червей и моллюсков и в сперматогенных клетках насекомых. Эдуард Страсбургер исследовал митотическое деление в клетках зелёной водоросли спирогиры, в материнских клетках пыльцы лука и в материнских споровых клетках плауна. Ссылаясь на работу Отто Бючли и основываясь на собственных исследованиях, Эдуард Страсбургер обратил внимание на единство процессов клеточного деления в растительных и животных клетках[16].
К концу 1878 — началу 1880 года появились подробные работы В. Шлейхера (о делении хрящевых клеток амфибий), В. Флемминга (о размножении клеток в разных тканях саламандры и её личинок), П. И. Перемежко (о делении клеток в эпидермисе личинок тритона). В своей работе в 1879 году Шлейхер предложил термин «кариокинез» для обозначения сложных процессов клеточного деления, подразумевая перемещения составных частей ядра[17]. Вальтер Флемминг впервые для обозначения непрямого деления клетки ввёл термин «митоз», который впоследствии стал общепринятым[12]. Также Флеммингу принадлежит окончательная формулировка определения митоза как циклического процесса, завершающегося разделением хромосом между дочерними клетками[18].
-
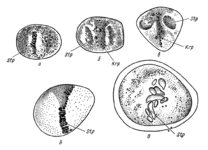 Деление клеток по Э. Руссову (1872)
Деление клеток по Э. Руссову (1872) -
 Деление клеток по Э. Страсбургеру (1875)
Деление клеток по Э. Страсбургеру (1875) -
 Деление клеток по В. Флеммингу (1882)
Деление клеток по В. Флеммингу (1882) -
 Деление клеток по Э. Б. Уилсону (1900)
Деление клеток по Э. Б. Уилсону (1900)
.png)
.png)
.png)

В 1880 году О. В. Баранецкий установил спиральное строение хромосом. В ходе дальнейших исследований были развиты представления о спирализации и деспирализации хромосом во время митотического цикла[18]. В начале 1900-х годов хромосомы были идентифицированы в качестве носителей наследственной информации, что в дальнейшем дало объяснение биологической роли митоза, заключающейся в образовании генетически идентичных дочерних клеток.
В 1970-х годах началась расшифровка и детальное изучение регуляторов митотического деления[19], благодаря серии экспериментов по слиянию клеток, находящихся на разных этапах клеточного цикла. В тех опытах, когда клетку в М-фазе объединяли с клеткой, находящейся в любой из стадий интерфазы (G1, S или G2), интерфазные клетки переходили в митотическое состояние (начиналась конденсация хромосом и распадалась ядерная мембрана)[20]. В итоге был сделан вывод, что в цитоплазме митотической клетки присутствует фактор (или факторы), стимулирующий митоз[21], или, иначе, М-стимулирующий фактор (МСФ, от англ. M-phase-promoting factor, MPF)[22].
Впервые «фактор стимуляции митоза» был открыт в зрелых неоплодотворенных яйцах шпорцевой лягушки, находящихся в М-фазе клеточного цикла. Цитоплазма такого яйца, инъецированная в ооцит, приводила к преждевременному переходу в М-фазу и к началу созревания ооцита (первоначально сокращение MPF означало Maturation promoting factor, что переводится как «фактор, способствующий созреванию»). В ходе дальнейших экспериментов были установлены универсальное значение и вместе с тем высокая степень консервативности «фактора стимуляции митоза»: экстракты, приготовленные из митотических клеток весьма разнообразных организмов (млекопитающих, морских ежей, моллюсков, дрожжей), при введении в ооциты шпорцевой лягушки переводили их в М-фазу[23].
В ходе последующих исследований выяснилось, что фактор, стимулирующий митоз, представляет собой гетеродимерный комплекс, состоящий из белка циклина и зависимой от циклина протеинкиназы. Циклин является регуляторным белком и обнаруживается у всех эукариот. Его концентрация периодически возрастает в течение клеточного цикла, достигая максимума в метафазе митоза. С началом анафазы наблюдается резкое сокращение концентрации циклина, вследствие его расщепления с помощью сложных белковых протеолитических комплексов — протеасом. Зависимая от циклина протеинкиназа представляет собой фермент (фосфорилазу), модифицирующий белки за счёт переноса фосфатной группы от АТФ на аминокислоты серин и треонин. Таким образом с установления роли и структуры основного регулятора митотического деления начались исследования тонких регуляторных механизмов митоза, которые продолжаются до настоящего времени.
Аппарат клеточного деления[править | править код]
Деление всех эукариотических клеток сопряжено с формированием специального аппарата клеточного деления. Активная роль в митотическом делении клеток зачастую отведена цитоскелетным структурам. Универсальным как для животных, так и для растительных клеток является двухполюсное митотическое веретено, состоящее из микротрубочек и связанных с ними белков[24]. Веретено деления обеспечивает строго одинаковое распределение хромосом между полюсами деления, в области которых в телофазе образуются ядра дочерних клеток.
Ещё одна не менее важная структура цитоскелета отвечает за разделение цитоплазмы (цитокинез) и, как следствие, за распределение клеточных органелл. В животных клетках за цитокинез отвечает сократимое кольцо из актиновых и миозиновых филаментов. В большинстве клеток высших растений из-за наличия жёсткой клеточной стенки цитокинез протекает с образованием клеточной пластинки в плоскости между двумя дочерними клетками. При этом область образования новой клеточной перегородки определяется заранее предпрофазным пояском из актиновых микрофиламентов, а поскольку актин участвует также в формировании клеточных септ у грибов, возможно, что он направляет цитокинез у всех эукариот[25].
Веретено деления[править | править код]

Формирование веретена деления начинается в профазе. В его образовании принимают участие полярные тельца (полюса) веретена и кинетохоры хромосом, и те и другие взаимодействуют с микротрубочками — биополимерами, состоящими из субъединиц тубулина. Главным центром организации микротрубочек (ЦОМТ) во многих эукариотических клетках является центросома — скопление аморфного фибриллярного материала, причём в большинстве животных клеток в состав центросом также входят пары центриолей[27]. Во время интерфазы ЦОМТ, как правило, располагающийся вблизи клеточного ядра, инициирует рост микротрубочек, расходящихся к периметру клетки и образующих цитоскелет. В S-фазе материал центросомы удваивается, а в профазе митоза начинается расхождение дочерних центросом. От них в свою очередь «отрастают» микротрубочки, которые удлиняются вплоть до соприкосновения друг с другом, после чего центросомы расходятся. Затем, в прометафазе, после разрушения ядерной мембраны, микротрубочки проникают в область клеточного ядра и взаимодействуют с хромосомами. Две дочерние центросомы теперь называют полюсами веретена[28].
По морфологии различают два типа митотического веретена: астральный (или конвергентный) и анастральный (дивергентный)[~ 1][30].
Астральный тип митотической фигуры, характерный для животных клеток, отличают благодаря небольшим зонам, на полюсах веретена, в которых сходятся (конвергируют) микротрубочки. Зачастую центросомы, располагающиеся в области полюсов астрального веретена, содержат центриоли. От полюсов деления также расходятся во всех направлениях радиальные микротрубочки, не входящие в состав веретена, а образующие звездчатые зоны — цитастеры.
Анастральный тип митотической фигуры отличается широкими полярными областями веретена, так называемыми полярными шапочками, в их состав не входят центриоли. Микротрубочки при этом расходятся широким фронтом (дивергируют) от всей зоны полярных шапочек. Этот тип митотической фигуры также отличает отсутствие цитастеров. Анастральный тип митотического веретена наиболее характерен для делящихся клеток высших растений, хотя иногда наблюдается и в некоторых клетках животных.
Микротрубочки[править | править код]
Микротрубочки — динамичные структуры, принимающие активное участие в построении веретена деления во время митоза. Химически они представляют собой биополимеры, состоящие из субъединиц белка тубулина. Количество микротрубочек в клетках различных организмов может значительно отличаться. В метафазе веретено деления в клетках высших животных и растений может содержать до нескольких тысяч микротрубочек, тогда как у некоторых грибов их всего около 40[28].
Митотические микротрубочки веретена деления «динамически нестабильны». Их «положительные» или «плюс-концы», расходящиеся во всех направлениях от центросом резко переходят от равномерного роста к стремительному укорочению, при котором часто деполимеризуется вся микротрубочка. Согласно этим данным, образование митотического веретена объясняется селективной (выборочной) стабилизацией микротрубочек, взаимодействующих в экваториальной области клетки с кинетохорами хромосом и с микротрубочками, идущими от противоположного полюса деления. Эта модель объясняет характерную двухполюсную фигуру митотического веретена[28].
Центромеры и кинетохоры[править | править код]
Центромеры — специализированные последовательности ДНК, необходимые для связывания с микротрубочками веретена деления и для последующего расхождения хромосом. В зависимости от локализации различают несколько типов центромер. Для голоцентрических центромер характерно образование связей с микротрубочками веретена по всей длине хромосомы (некоторые насекомые, нематоды, некоторые растения). В противоположность голоцентрическим моноцентрические центромеры служат для связи с микротрубочками в единственной области хромосомы[30].
В центромерной области обычно располагаются кинетохоры хромосом — сложные белковые комплексы, морфологически очень сходные по своей структуре для различных групп эукариот, как, например, для диатомовых водорослей, так и для человека[31]. Обычно на каждую хроматиду (хромосому) приходится по одному кинетохору. На электронных микрофотографиях кинетохор обычно выглядит как пластинчатая трёхслойная структура[32]. Порядок слоев следующий: внутренний плотный слой, примыкающий к телу хромосомы; средний рыхлый слой; внешний плотный слой, от которого отходит множество фибрилл, образуя т. н. фиброзную корону кинетохора.
К основным функциям кинетохора относят: закрепление микротрубочек веретена деления, обеспечение движения хромосом во время митоза при участии микротрубочек, связывание между собой сестринских хроматид и регуляцию их последующего разделения в анафазе митоза[33]. Минимально достаточно одной микротрубочки (например, для дрожжей) ассоциированной с кинетохором, чтобы обеспечить движение хромосомы. Однако с одним кинетохором могут быть связаны целые пучки, состоящие из 20—40 микротрубочек (например, у высших растений или человека), чтобы обеспечить расхождение хромосом к полюсам клетки[32][33].
Продолжительность митоза[править | править код]
.svg)
Собственно митоз зачастую протекает сравнительно быстро. Средняя продолжительность составляет 1—2 часа[3][6], что занимает всего около 10 % времени клеточного цикла. К примеру, у делящихся клеток меристемы корней интерфаза составляет 16—30 часов, а митоз длится всего 1—3 часа. Для эпителиальных клеток кишечника мыши интерфазный период составляет порядка 20—22 часов, а митоз продолжается в течение 1 часа[34]. В клетках животных митоз обычно протекает быстрее и длится в среднем 30—60 минут, в то время как в растительных клетках средняя продолжительность митоза составляет 2—3 часа[7]. Известны исключения с противоположными показателями. К примеру, в животных клетках продолжительность митоза может достигать 3,8 часов (эпидермис мыши). Или же встречаются растительные объекты с длительностью митоза в 5 минут (Chilomonas)[35]. Наиболее интенсивно митоз протекает в эмбриональных клетках (10—40 минут в дробящихся яйцеклетках).
Длительность митоза находится в зависимости от целого ряда факторов: размеров делящейся клетки, её плоидности, числа ядер. Частота клеточных делений также зависит от степени дифференцировки клеток и специфики выполняемых функций. Так, нейроны или клетки скелетной мышцы человека не делятся совсем; клетки печени обычно делятся раз в один или два года, а некоторые эпителиальные клетки кишечника делятся чаще, чем 2 раза в сутки[36].
Темп клеточного деления зависит также от условий окружающей среды, в частности, от температуры. Повышение температуры окружающей среды в физиологических пределах повышает скорость митоза, что может быть объяснено обычной закономерностью кинетики химических реакций[37].
Фазы митоза[править | править код]
Фаза клеточного цикла, соответствующая делению клетки, называется М-фазой (от слова «митоз»). М-фазу условно подразделяют на шесть стадий, постепенно и непрерывно переходящих одна в другую[27][34]. Первые пять — профаза, прометафаза (метакинез), метафаза, анафаза и телофаза (или цитотомия) — составляют митоз[~ 2], а берущий своё начало в анафазе процесс разделения цитоплазмы клетки, или цитокинез, протекает вплоть до завершения митотического цикла и, как правило, рассматривается в составе телофазы.
Длительность отдельных стадий различна и варьируется в зависимости от типа ткани, физиологического состояния организма, внешних факторов. Наиболее продолжительны стадии, сопряженные с процессами внутриклеточного синтеза: профаза (2—270 минут) и телофаза (1,5—140 минут). Наиболее быстротечны фазы митоза, в ходе которых происходит движение хромосом: метафаза (0,3—175 минут) и анафаза (0,3—122 минуты). Непосредственно процесс расхождения хромосом к полюсам обычно не превышает 10 минут[39].
Препрофаза[править | править код]

1) Клетка с крупной центральной вакуолью и ядром, оттеснённым на периферию клетки.
2) Полоски цитоплазмы рассекают вакуоль, обеспечивая перемещение ядра к центру клетки.
3) и 4) Полоски цитоплазмы объединяются в плоскости деления клетки с образованием фрагмосомы.
5) На этапе цитокинеза в области фрагмосомы образуется фрагмопласт и новая клеточная стенка

1) и 2) Оформление препрофазного кольца.
3) Нуклеация микротрубочек вокруг ядра.
4) Препрофазное кольцо начинает исчезать.
5) Микротрубочки вокруг ядра образуют «профазное веретено».
6) Ядерная мембрана распадается и микротрубочки веретена направляются к хромосомам.
Условные обозначения: N — ядро; V — вакуоль; PPB — препрофазное кольцо; MTN — начало скопления микротрубочек; NEB — распад ядерной оболочки; микротрубочки окрашены зелёным цветом
Препрофаза — редко используемый термин[40] для обозначения дополнительной стадии митоза растительной клетки. К основным событиям препрофазы можно отнести: оформление препрофазного кольца, образование фрагмосомы и начало нуклеации микротрубочек вокруг клеточного ядра. Несмотря на наличие термина «препрофаза», перечисленные события чаще рассматриваются в составе фазы G2[40][41][42] или в составе профазы[40][43].
В клетках, богатых вакуолями, в ходе препрофазы образуется фрагмосома — одна из структур, определяющих плоскость деления растительной клетки. Фрагмосома представляет собой слой цитоплазмы, пересекающий вакуоль в плоскости клеточного деления[44]. Ядро в клетках с крупной центральной вакуолью обычно располагается на периферии. Во время препрофазы оно перемещается в область фрагмосомы. В ходе перемещения ядра вакуоль рассекается полосками цитоплазмы, содержащими элементы цитоскелета. Во фрагмосоме также образуется митотическое веретено. Во время цитокинеза в области фрагмосомы формируется фрагмопласт и новая клеточная стенка.
Одновременно с фрагмосомой образуется препрофазное кольцо, и обе структуры располагаются в одной плоскости[45]. Препрофазное кольцо — это кольцеобразное скопление микротрубочек и актиновых филаментов вблизи клеточной мембраны в плоскости деления растительной клетки. Ядро располагается в центре препрофазного кольца и связано с ним радиально расходящимися микротрубочками. Внешне эта структура напоминает колесо с ободом и спицами из микротрубочек и актиновых филаментов, а также с ядром на месте втулки[45]. Структура кольца также обогащена элементами ЭПР и везикулами аппарата Гольджи.
Препрофазное кольцо формируется перед профазой митоза. После начала профазы микротрубочки кольца деполимеризуются и далее участвуют в образовании веретена деления. Функции препрофазного кольца пока не ясны. Однако подмечено, что цитокинез растительной клетки происходит в плоскости определённой положением препрофазного кольца[40]. При симметричном делении кольцо формируется в середине, а при асимметрическом делении образуется ближе к одному концу клетки[45].
Профаза[править | править код]
К основным событиям профазы относят конденсацию хромосом внутри ядра и образование веретена деления в цитоплазме клетки[46]. Распад ядрышка в профазе является характерной, но необязательной для всех клеток особенностью[47].
Условно за начало профазы принимается момент возникновения микроскопически видимых хромосом вследствие конденсации внутриядерного хроматина. Уплотнение хромосом происходит за счёт многоуровневой спирализации ДНК. Эти изменения сопровождаются повышением активности фосфорилаз, модифицирующих гистоны, непосредственно участвующие в компоновке ДНК. Как следствие, резко снижается транскрипционная активность хроматина, инактивируются ядрышковые гены, большая часть ядрышковых белков диссоциирует. Конденсирующиеся сестринские хроматиды в ранней профазе остаются спаренными по всей своей длине с помощью белков-когезинов, однако к началу прометафазы связь между хроматидами сохраняется лишь в области центромер. К поздней профазе на каждой центромере сестринских хроматид формируются зрелые кинетохоры, необходимые хромосомам для присоединения к микротрубочкам веретена деления в прометафазе[48].

Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое веретено — одна из главных структур аппарата клеточного деления, ответственная за распределение хромосом между дочерними клетками. В образовании веретена деления у всех эукариотических клеток принимают участие полярные тельца (центросомы), микротрубочки и кинетохоры хромосом[30].
С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических свойств микротрубочек. Время полужизни средней микротрубочки уменьшается примерно в 20 раз от 5 минут (в интерфазе) до 15 секунд[28][48]. Однако скорость их роста увеличивается примерно в 2 раза по сравнению с теми же интерфазными микротрубочками[48]. Полимеризующиеся плюс-концы («+»-концы) являются «динамически нестабильными» и резко переходят от равномерного роста к быстрому укорочению, при котором часто деполимеризуется вся микротрубочка[28]. Для правильного функционирования митотического веретена необходим определённый баланс между процессами сборки и деполимеризации микротрубочек, так как ни стабилизированные, ни деполимеризованные микротрубочки веретена не в состоянии перемещать хромосомы[~ 3].
Наряду с наблюдаемыми изменениями динамических свойств микротрубочек, слагающих нити веретена, в профазе закладываются полюса деления. Реплицированные в S-фазе центросомы расходятся в противоположных направлениях за счёт взаимодействия полюсных микротрубочек, растущих навстречу друг другу. Своими минус-концами («−»-концами) микротрубочки погружены в аморфное вещество центросом, а процессы полимеризации протекают со стороны плюс-концов, обращенных к экваториальной плоскости клетки. При этом вероятный механизм расхождения полюсов объясняется следующим образом: динеино-подобные белки ориентируют в параллельном направлении полимеризующиеся плюс-концы полюсных микротрубочек, а кинезино-подобные белки в свою очередь расталкивают их в направлении к полюсам деления[50].
Параллельно конденсации хромосом и формированию митотического веретена, во время профазы происходит фрагментация эндоплазматического ретикулума, который распадается на мелкие вакуоли, расходящиеся затем к периферии клетки. Одновременно рибосомы теряют связи с мембранами ЭПР. Цистерны аппарата Гольджи также меняют свою околоядерную локализацию, распадаясь на отдельные диктиосомы, без особого порядка распределенные в цитоплазме[51].
Прометафаза[править | править код]

Окончание профазы и наступление прометафазы, как правило, знаменуется распадом ядерной мембраны[46]. Целый ряд белков ламины фосфорилируется, вследствие чего ядерная оболочка фрагментируется на мелкие вакуоли, а поровые комплексы исчезают[52]. После разрушения ядерной мембраны хромосомы без особого порядка располагаются в области ядра. Однако вскоре все они приходят в движение.
В прометафазе наблюдается интенсивное, но беспорядочное перемещение хромосом. Поначалу отдельные хромосомы стремительно дрейфуют к ближайшему полюсу митотического веретена со скоростью, достигающей 25 мкм/мин[52]. Вблизи полюсов деления повышается вероятность взаимодействия новосинтезированных плюс-концов микротрубочек веретена с кинетохорами хромосом[52][53]. В результате такого взаимодействия кинетохорные микротрубочки (связанные с кинетохором) стабилизируются от спонтанной деполимеризации, а их рост отчасти обеспечивает отдаление соединенной с ними хромосомы в направлении от полюса к экваториальной плоскости веретена. С другой стороны хромосому настигают тяжи микротрубочек, идущие от противоположного полюса митотического веретена. Взаимодействуя с кинетохором, они также участвуют в движении хромосомы. В результате сестринские хроматиды оказываются связанными с противоположными полюсами веретена[49]. Усилие, развиваемое микротрубочками от разных полюсов, не только стабилизирует взаимодействие этих микротрубочек с кинетохорами, но также, в конечном счёте, приводит каждую хромосому в плоскость метафазной пластинки[54].
В клетках млекопитающих прометафаза протекает, как правило, в течение 10—20 минут[53]. В нейробластах кузнечика эта стадия занимает всего 4 минуты, а в эндосперме Haemanthus и в фибробластах тритона — около 30 минут[55]. В дрожжевых клетках невозможно чётко разграничить стадии профазы и прометафазы по причине сохранения ядерной оболочки в процессе деления. Аналогичным образом, частичное или более позднее разрушение ядерной мембраны затрудняет разграничение стадий профазы и прометафазы в клетках Drosophila и C. elegans. В подобных случаях для описания всех ранних событий митотического деления используется обобщающий термин «профаза»[46].
Метафаза[править | править код]

В завершении прометафазы хромосомы располагаются в экваториальной плоскости веретена (а не всей клетки[56]) примерно на равном расстоянии от обоих полюсов деления, образуя метафазную (экваториальную) пластинку. Морфология метафазной пластинки в клетках животных, как правило, отличается упорядоченным расположением хромосом: центромерные участки обращены к центру веретена, а плечи — к периферии клетки (фигура «материнской звезды»). В растительных клетках хромосомы зачастую лежат в экваториальной плоскости веретена без строгого порядка[57][58]. В дрожжевых клетках хромосомы тоже не выстраиваются в экваториальной плоскости, а располагаются произвольно вдоль волокон веретена деления[46].
Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием. Все это время хромосомы удерживаются в экваториальной плоскости веретена за счёт сбалансированных сил натяжения кинетохорных микротрубочек, совершая колебательные движения с незначительной амплитудой в плоскости метафазной пластинки[59].
В метафазе, также как и в течение других фаз митоза, продолжается активное обновление микротрубочек веретена путём интенсивной сборки и деполимеризации молекул тубулина. Несмотря на некоторую стабилизацию пучков кинетохорных микротрубочек, происходит постоянная переборка межполюсных микротрубочек, численность которых в метафазе достигает максимума[57].
К окончанию метафазы наблюдается чёткое обособление сестринских хроматид, соединение между которыми сохраняется лишь в центромерных участках. Плечи хроматид располагаются параллельно друг другу, и становится отчетливо заметной разделяющая их щель[57].
Анафаза[править | править код]

Анафаза — самая короткая стадия митоза, которая начинается внезапным разделением и последующим расхождением сестринских хроматид в направлении противоположных полюсов клетки[60]. Хроматиды расходятся с равномерной скоростью достигающей 0,5—2 мкм/мин[3][61] (0,2—5 мкм/мин[62]), при этом они часто принимают V-образную форму. Их движение обусловлено воздействием значительных сил, оценочно 10−5 дин на хромосому, что в 10 000 раз превышает усилие, необходимое для простого продвижения хромосомы через цитоплазму с наблюдаемой скоростью[63].
Как правило, расхождение хромосом в анафазе состоит из двух относительно независимых процессов, называемых анафазой А и анафазой В.
Анафаза А характеризуется расхождением сестринских хроматид к противоположным полюсам деления клетки[46]. За их движение при этом отвечают те же силы, что ранее удерживали хромосомы в плоскости метафазной пластинки. Процесс расхождения хроматид сопровождается сокращением длины деполимеризующихся кинетохорных микротрубочек. Причем их распад наблюдается преимущественно (на 80 %[64]) в области кинетохоров, со стороны плюс-концов (ранее, с начала профазы и вплоть до начала анафазы, на плюс-концах преобладали процессы сборки субъединиц тубулина)[63]. Вероятно, деполимеризация микротрубочек у кинетохоров либо в области полюсов деления является необходимым условием для перемещения сестринских хроматид, так как их движение прекращается при добавлении таксола или тяжёлой воды (D2O), оказывающих стабилизирующее воздействие на микротрубочки. Механизм, лежащий в основе расхождения хромосом в анафазе А, пока остается неизвестным[~ 4][63].
Во время анафазы В расходятся сами полюса деления клетки[46], и, в отличие от анафазы А, этот процесс происходит за счёт сборки полюсных микротрубочек со стороны плюс-концов. Полимеризующиеся антипараллельные нити веретена при взаимодействии отчасти и создают расталкивающее полюса усилие. Величина относительного перемещения полюсов при этом, также как и степень перекрывания полюсных микротрубочек в экваториальной зоне клетки, сильно варьирует у особей разных видов[65]. Помимо расталкивающих сил, на полюса деления воздействуют тянущие силы со стороны астральных микротрубочек, которые создаются в результате взаимодействия с динеино-подобными белками на плазматической мембране клетки[66].
Последовательность, продолжительность и относительный вклад каждого из двух процессов, слагающих анафазу, могут быть крайне различны. Так, в клетках млекопитающих анафаза В начинается сразу вслед за началом расхождения хроматид к противоположным полюсам и продолжается вплоть до удлинения митотического веретена в 1,5—2 раза по сравнению с метафазным. В некоторых других клетках (например, дрожжевых) анафаза В начинается только после того как хроматиды достигают полюсов деления. У некоторых простейших в процессе анафазы В веретено удлиняется в 15 раз по сравнению с метафазным[60]. В растительных клетках анафаза В отсутствует[66].
Телофаза[править | править код]

Телофаза (от греч. τέλος — конец) рассматривается как заключительная стадия митоза; за её начало принимается момент остановки разделённых сестринских хроматид у противоположных полюсов деления клетки[66]. В ранней телофазе наблюдается деконденсация хромосом и, следовательно, увеличение их в объёме. Вблизи сгруппированных индивидуальных хромосом начинается слияние мембранных пузырьков, что дает начало реконструкции ядерной оболочки. Материалом для построения мембран новообразованных дочерних ядер служат фрагменты изначально распавшейся ядерной мембраны материнской клетки, а также элементы эндоплазматического ретикулума[67]. При этом отдельные пузырьки связываются с поверхностью хромосом и сливаются воедино. Постепенно восстанавливается наружная и внутренняя ядерные мембраны, восстанавливаются ядерная ламина и ядерные поры. В процессе восстановления ядерной оболочки дискретные мембранные пузырьки, вероятно, соединяются с поверхностью хромосом без распознавания специфических последовательностей нуклеотидов, так как в результате проведенных экспериментов было выявлено, что восстановление ядерной мембраны происходит вокруг молекул ДНК, заимствованных у любого организма, даже у бактериального вируса[68]. Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и становятся различимыми ядрышки.
Параллельно с процессами образования ядер дочерних клеток в телофазе начинается и заканчивается разборка микротрубочек веретена деления. Деполимеризация протекает в направлении от полюсов деления к экваториальной плоскости клетки, от минус-концов к плюс-концам. При этом дольше всего сохраняются микротрубочки в средней части веретена деления, которые образуют остаточное тельце Флемминга[69].
Цитокинез[править | править код]

Окончание телофазы преимущественно совпадает с разделением тела материнской клетки — цитокинезом (цитотомией)[70][71]. При этом образуются две или более дочерние клетки. Процессы, ведущие к разделению цитоплазмы, берут своё начало ещё в середине анафазы и могут продолжаться после завершения телофазы. Митоз не всегда сопровождается разделением цитоплазмы, поэтому цитокинез не классифицируется в качестве отдельной фазы митотического деления и обычно рассматривается в составе телофазы[~ 5].
Различают два основных типа цитокинеза: деление поперечной перетяжкой клетки (наиболее характерно для клеток животных) и деление путём образования клеточной пластинки (свойственно растениям в связи с наличием жёсткой клеточной стенки). Плоскость деления клетки детерминируется положением митотического веретена и проходит под прямым углом к длинной оси веретена[72].
При делении поперечной перетяжкой клетки место разделения цитоплазмы закладывается предварительно ещё в период анафазы, когда в плоскости метафазной пластинки под мембраной клетки возникает сократительное кольцо из актиновых и миозиновых филаментов. В дальнейшем, вследствие активности сократительного кольца, образуется борозда деления, которая постепенно углубляется вплоть до полного разделения клетки. По окончании цитокинеза сократимое кольцо полностью распадается, а плазматическая мембрана стягивается вокруг остаточного тельца Флемминга, состоящего из скопления остатков двух групп полюсных микротрубочек, тесно упакованных вместе с материалом плотного матрикса[73].
Деление путём образования клеточной пластинки начинается с перемещения мелких ограниченных мембраной пузырьков по направлению к экваториальной плоскости клетки. Здесь они сливаются, образуя дисковидную, окружённую мембраной структуру — раннюю клеточную пластинку. Мелкие пузырьки происходят в основном из аппарата Гольджи и перемещаются к экваториальной плоскости вдоль остаточных полюсных микротрубочек веретена деления, образующих цилиндрическую структуру, называемую фрагмопластом. По мере расширения клеточной пластинки микротрубочки раннего фрагмопласта попутно перемещаются к периферии клетки, где за счёт новых мембранных пузырьков продолжается рост клеточной пластинки вплоть до её окончательного слияния с мембраной материнской клетки. После окончательного разделения дочерних клеток в клеточной пластинке откладываются микрофибриллы целлюлозы, завершая образование жёсткой клеточной стенки[74].
Регуляция митоза[править | править код]
Основными регулято́рными механизмами митоза являются процессы фосфорилирования и протеолиза[75]. Обратимые реакции фосфорилирования и дефосфорилирования обеспечивают протекание обратимых событий митоза, таких как сборка/распад веретена деления или распад/восстановление ядерной оболочки. Протеолиз лежит в основе необратимых событий митоза, таких как разделение сестринских хроматид в анафазе или разрушение митотических циклинов на поздних стадиях митоза.
Контрольные точки[править | править код]
.svg)
Рассматривая вопрос регуляции митоза, можно условно выделить два периода митотического деления: от начала профазы до анафазы, и далее, от анафазы до конца телофазы[77]. Каждый из двух обозначенных периодов начинается с прохождения контрольной точки клеточного цикла.
Первой контрольной точкой является переход из фазы G2 к M-фазе. Главным условием для преодоления контрольной точки G2/M является завершённая репликация ДНК: старт митотического деления блокируется у большинства эукариот при повреждениях или неоконченной репликации ДНК. События от начала профазы и до окончания метафазы инициируются и протекают при участии белковых комплексов, состоящих из митотических циклинов и циклин-зависимых киназ (англ. M-Cdk).
Вторая контрольная точка служит разделительным барьером на границе метафазы и анафазы. На этом этапе критичным показателем является состояние веретена деления: вступление в анафазу у всех эукариот блокируется в случае наличия дефектов веретена. Ключевым активатором событий анафазы является убиквитинлигаза APCCdc20[76].
Основные регуляторы митоза[править | править код]
Циклин-киназы[править | править код]

1 — неактивная циклин-зависимая протеинкиназа;
2 — циклин;
3 — активный циклин-киназный комплекс

Ключевыми активаторами митоза, обеспечивающими инициацию событий профазы—метафазы, являются циклин-киназные комплексы (англ. M-Cdk). Эти комплексы представляют собой гетеродимеры, состоящие из двух субъединиц: регуляторной — митотического циклина (англ. M cyclin) и каталитической — циклин-зависимой киназы (англ. Cdk — cyclin-dependent kinase).
В регуляцию митоза у всех эукариот вовлечена циклин-зависимая киназа Cdk1[79], которая представляет собой фермент (фосфорилазу), модифицирующий белки за счёт переноса фосфатной группы от АТФ на аминокислоты серин и треонин. Концентрация Cdk1 постоянна на протяжении всего клеточного цикла[80], поэтому активность циклин-зависимой киназы в процессе митоза зависит главным образом от её соединения с митотическим циклином. Концентрация митотических циклинов увеличивается по мере приближения к митозу и достигает максимума в метафазе. Различным таксонам свойственны различные митотические циклины. Так, у почкующихся дрожжей в регуляцию митоза вовлечены четыре циклина Clb1, 2, 3 и 4; у дрозофилы — циклины A, B, B3; у позвоночных — циклин B[81].
Регуляторы активности циклин-киназ[править | править код]

Накопление митотических циклинов начинается ещё на стадии G2. Увеличение концентрации циклинов обеспечивается за счёт транскрипции соответствующих им генов[83]. Новосинтезированные циклины сразу объединяются с неактивной киназой Cdk1. Однако образующиеся при этом циклин-киназные комплексы сохраняются в неактивном состоянии до момента активации митоза. Сдерживание активности комплексов M-Cdk1 на протяжении фазы G2 осуществляется благодаря ингибирующему фосфорилированию молекулы Cdk1[84]. За ингибирование Cdk1 ответственна группа протеинкиназ семейства Wee1[81][83]. В итоге, к началу митоза в клетке накапливается значительное количество неактивных комплексов M-Cdk1.
Собственно начало профазы на молекулярном уровне знаменуется резкой активацией киназных комплексов M-Cdk1. В основе скачкообразного увеличения активности M-Cdk1 лежат как минимум два взаимосвязанных события. Во-первых, к началу профазы приурочена активация фосфатаз семейства Cdc25, которые освобождают комплекс M-Cdk1 от ингибирующих фосфатных групп. Во-вторых, активированные таким образом киназы M-Cdk1 включаются в цепочку положительной обратной связи: путём фосфорилирования они активируют собственных активаторов семейства Cdc25 и ингибируют собственных ингибиторов семейства Wee1. В итоге, в начале профазы наблюдается взаимосвязанное увеличение активности фосфатаз семейства Cdc25 и циклин-киназ M-Cdk1 на фоне параллельного снижения активности ингибиторов семейства Wee1. Таким образом, в основу активации митоза заложен принцип положительной обратной связи. Но, несмотря на то, что уже известно об инициирующих механизмах митоза, остаётся до сих пор неясно, какой именно стимул изначально активирует Cdc25 или Cdk1, тем самым обеспечивая запуск цепочки положительной обратной связи[~ 6][83][86].
Polo- и aurora-подобные киназы[править | править код]
Помимо циклин-зависимых киназ в регуляцию митотических событий вовлечены как минимум ещё два типа киназ: polo-подобные киназы и киназы семейства aurora. Polo-подобные киназы (англ. polo-like kinase, Plk) представляют собой серин-треониновые протеинкиназы, активирующиеся на начальных и инактивирующиеся на поздних стадиях митоза или в начале фазы G1. Эти киназы вовлечены в различные митотические процессы: сборку веретена деления, функционирование кинетохора, цитокинез[87]. Киназы семейства aurora также относятся к группе серин-треониновых протеинкиназ. У многоклеточных выделяются два основных представителя этого семейства: aurora A и aurora B. Киназа aurora A участвует в регуляции функционирования центросом и митотического веретена. Киназа aurora B участвует в регуляции процессов конденсации и разделения сестринских хроматид, а также обеспечивает присоединение кинетохоров к микротрубочкам веретена деления[88].
Активатор анафазы APCCdc20[править | править код]
.svg)
Комплекс, стимулирующий анафазу (англ. anaphase-promoting complex, APC), также называемый циклосома, представляет собой крупное белковое соединение, которому отводится решающая роль в активации анафазы. Функционально комплекс стимуляции анафазы представляет собой убиквитинлигазу и катализирует реакции присоединения молекул убиквитина к различным целевым белкам, которые в итоге подвергаются протеолизу[90].
В структуре комплекса стимуляции анафазы выделяется порядка 11—13 субъединиц. Ядро комплекса составляют: субъединица куллина (Apc2) и RING-домен (Apc11), к которому присоединяется убиквитин-конъюгирующий фермент (E2). Функционирование комплекса регулируется за счёт присоединения активирующей субъединицы в нужный момент клеточного цикла[89].
Белок Cdc20 (англ. cell division cycle protein 20 — «белок клеточного цикла 20») активирует комплекс APC при переходе делящейся клетки из метафазы в анафазу. Происходит это следующим образом. На стадии метафазы циклин-киназный комплекс M-Cdk путём фосфорилирования трансформирует ядро комплекса APC. В результате указанного конформационного изменения повышается вероятность присоединения активатора Cdc20. В итоге, активированный комплекс APCCdc20 обретает убиквитин-лигазную активность и убиквитинирует свои главные цели — секьюрин и митотические циклины[89].
Секьюрин (одна из главных мишеней APCCdc20) представляет собой ингибирующий белок, сдерживающий в неактивном состоянии фермент сепаразу. Вследствие реакции убиквитинирования секьюрин разрушается, а высвободившаяся при этом сепараза разрушает когезин. После деградации когезина, обеспечивающего сцепление сестринских хроматид, происходит разделение и расхождение хромосом к полюсам деления клетки[91].
Убиквитинирование и, как следствие, разрушение митотических циклинов (ещё одной важной мишени APCCdc20) запускает цепочку отрицательной обратной связи. Выглядит это следующим образом. Циклин-киназный комплекс M-Cdk активирует убиквитин-лигазный комплекс APCCdc20, который целенаправленно разрушает митотические циклины, что ведёт к деградации циклин-киназного комплекса M-Cdk, то есть цепочка реакций приводит к разрушению изначального активатора этой цепочки. Но поскольку активность APCCdc20 зависит от комплекса M-Cdk, инактивация циклин-киназы M-Cdk приводит к инактивации APCCdc20. В итоге APCCdc20 деактивируется к концу митоза[89].
Митотический кроссинговер[править | править код]

Митотический кроссинговер — процесс обмена участками гомологичных хромосом в процессе митотического деления. Относительно редкий тип генетической рекомбинации в соматических клетках, по причине отсутствия нормального механизма конъюгации хромосом[92][93]. Частота митотического кроссинговера составляет не более одного раза на миллион клеточных делений[94] (1,3±0,1 на 106 клеточных делений[95]). У некоторых диплоидных грибов частота митотической рекомбинации может достигать 1—10 % от частоты мейотического кроссинговера[96]. Воздействие радиации или химических реагентов может повысить частоту митотической рекомбинации. Некоторые исследователи предполагают сходство механизмов мейотического и митотического кроссинговера[95].
Первые данные о существовании митотической рекомбинации были получены генетиком Куртом Штерном в 1936 году. Учёный проводил исследования на плодовых мушках и обратил внимание на локальное проявление рецессивных признаков у гетерозиготных особей. То есть у мух с нормальным внешним покровом появлялись участки ткани с жёлтым окрасом или с «опалёнными» щетинками. Однако оба признака кодировались генами, локализованными в пределах одной хромосомы, и у гетерозиготных особей не должны были проявляться. Особо любопытны оказались случаи «двойных пятен», при которых проявлялись сразу оба рецессивных признака, причём у особей как женского, так и мужского пола. В итоге, на основе полученных данных был сделан вывод о существовании митотической рекомбинации в соматических клетках[94][95].
Патология митоза[править | править код]
Патология митоза развивается при нарушении нормального течения митотического деления и зачастую приводит к возникновению клеток с несбалансированными кариотипами, следовательно, ведёт к развитию мутаций и анеуплоидии. Также в результате развития отдельных форм патологии наблюдаются хромосомные аберрации. Незавершённые митозы, прекращающиеся по причине дезорганизации или разрушения митотического аппарата, приводят к образованию полиплоидных клеток. Полиплоидия и формирование дву- и многоядерных клеток возникают в случае нарушений механизмов цитокинеза. При значительных последствиях патологии митоза возможна гибель клетки.
В нормальных тканях патология встречается в незначительных количествах. Например, в эпидермисе мышей встречается около 0,3 % патологических митозов; в эпителии гортани и матки человека — около 2 %. Патологические митозы часто наблюдаются при канцерогенезе, при различных экстремальных воздействиях, при лучевой болезни или вирусной инфекции[~ 7], при раке и предраковых гиперплазиях[~ 8]. Частота патологических митозов также увеличивается с возрастом[99].
Условно различают патологию митоза функционального и органического типа. К функциональным нарушениям относят, например, гипореактивность вступающих в митоз клеток — снижение реакции на физиологические регуляторы, определяющие интенсивность пролиферации нормальных клеток. Органические нарушения возникают при повреждении структур, участвующих в митотическом делении (хромосомы, митотический аппарат, клеточная поверхность), а также при нарушении процессов, связанных с этими структурами (репликация ДНК, образование веретена деления, движение хромосом, цитокинез)[99].
Классификация и общая характеристика различных форм патологий митоза[править | править код]
На основании морфологических признаков и цитохимических нарушений митотического процесса выделяют три основных группы патологий митоза: патология, связанная с повреждением хромосом; патология, связанная с повреждением митотического аппарата; нарушение цитокинеза[100].
I. Патология митоза, связанная с повреждением хромосом[править | править код]
1) Задержка митоза в профазе наблюдается при нарушениях репликации ДНК.
2) Нарушение спирализации и деспирализации хромосом прослеживается в результате действия на делящуюся клетку различными митотическими ядами. Например, воздействие колхицина приводит к гиперспирализации хромосом, которые приобретают укороченную и утолщенную форму[100].
3) Раннее (преждевременное) разделение хроматид в профазе (в норме разделение хроматид происходит на рубеже перехода метафазы в анафазу). Обозначенная патология наблюдается, к примеру, при изменении осмотического давления в фибробластах кролика в культуре ткани или же при воздействии канцерогенов (бензпирена, метилхолантрена) на мышиные фибробласты[100].


4) Фрагментация и пульверизация хромосом возникает в опухолевых клетках, при вирусной инфекции, в результате воздействия на нормальные клетки ионизирующего излучения или мутагенов. Фрагменты могут быть одиночными, парными и множественными. Те из них, которые лишены центромерного участка, не участвуют в метакинезе, и, соответственно, не расходятся к полюсам деления в анафазе. При массовой фрагментации хромосом (пульверизация) большинство фрагментов также беспорядочно рассеиваются в цитоплазме и не участвуют в метакинезе[101].
В итоге часть фрагментов хромосом может попасть в одно из дочерних ядер, либо резорбироваться, либо образовать обособленное микроядро. Также отдельные фрагменты обладают способностью воссоединяться своими концами, причём подобные воссоединения носят случайный характер и приводят к хромосомным аберрациям[102].
5) Хромосомные и хроматидные мосты являются следствием фрагментации хромосом. При воссоединении фрагментов содержащих центромер образуется дицентрическая хромосома, которая в ходе анафазы растягивается между противоположными полюсами деления, образуя мост. Хромосомный (обычно двойной) мост возникает в результате воссоединения фрагментов хромосом, каждый из которых образован двумя хроматидами с центромерой. Хроматидный (обычно одиночный) мост возникает в результате воссоединения двух фрагментов отдельных хроматид с центромерой[103].
К концу анафазы — в начале телофазы мосты обычно быстро рвутся в результате чрезмерного растягивания дицентрических фрагментов хромосом. Образование мостов приводит к генотипической разнородности дочерних клеток, а также нарушает течение завершающих стадий деления и задерживает цитокинез[103].
6) Отставание хромосом в метакинезе и при расхождении к полюсам возникает при повреждении хромосом в области кинетохора. Поврежденные хромосомы пассивно «дрейфуют» в цитоплазме и в итоге либо разрушаются и элиминируются из клетки, либо случайным образом попадают в одно из дочерних ядер, либо образуют отдельное микроядро. Отставание хромосом наблюдалось в культурах ткани опухолевых клеток, а также в экспериментах, в ходе которых кинетохоры хромосом облучались микропучком ультрафиолетовых лучей[104].
7) Образование микроядер происходит вследствие фрагментации или отставания отдельных хромосом, вокруг которых в телофазе формируется ядерная оболочка, параллельно образованию оболочки вокруг основных дочерних ядер. Новообразованные микроядра либо сохраняются в клетке в течение всего дальнейшего клеточного цикла вплоть до очередного деления, либо подвергаются пикнозу, разрушаются и выводятся из клетки[104].
8) При нерасхождении хромосом сестринские хроматиды не разъединяются с началом анафазы и вместе отходят к одному из полюсов, что приводит к анеуплоидии[105].
9) Набухание и слипание хромосом наблюдается в опухолевых клетках и при воздействии токсических доз различных митотических ядов. Вследствие набухания хромосомы теряют свои нормальные очертания и слипаются, превращаясь в комковатые массы. Расхождения хромосом не происходит и клетки в таком состоянии зачастую погибают[105].
II. Патология митоза, связанная с повреждением митотического аппарата[править | править код]
1) Задержка митоза в метафазе характерна для всей группы патологий митоза, связанных с повреждением митотического аппарата.
2) Колхициновый митоз или к-митоз — одна из форм патологии митоза, связанная с повреждением митотического аппарата вследствие воздействия статмокинетических ядов (колхицина, колцемида, винбластина, винкристина, аценафтена, нокодазол, метанола и др.)[106]. В результате воздействия статмокинетических ядов митоз задерживается на стадии метафазы в связи с дезорганизацией различных компонентов митотического веретена деления — центриолей, микротрубочек, кинетохоров. Повреждения также затрагивают клеточное ядро, плазмалемму, различные внутриклеточные органоиды (митохондрии, хлоропласты, аппарат Гольджи). Действие статмокинетических ядов усиливает спирализацию хромосом, что приводит к их укорочению и утолщению, а порой доводит до набухания и слипания хромосом. Как следствие, происходят хромосомные аберрации, образуются микроядра в результате фрагментации или отставания хромосом, развивается анеуплоидия[107].
Исход к-митоза зависит от дозы и времени воздействия статмокинетического яда на делящуюся клетку. При токсических дозах наблюдается пикноз ядра и гибель клетки. Значительные отравления приводят к полиплоидизации. Воздействие небольших доз обратимо. В течение нескольких часов может восстановиться митотический аппарат и продолжиться митотическое деление[107].
3) Рассеивание хромосом в метафазе происходит в результате повреждения или полной дезорганизации митотического аппарата.

4) Многополюсный митоз связан с аномалией репродукции центриолей, что ведет к формированию дополнительных полюсов и веретен деления. В итоге хромосомы распределяются неравномерно между дочерними ядрами, что, в свою очередь, ведет к образованию анеуплоидных клеток с несбалансированным набором хромосом[108].
5) Моноцентрический митоз связан с нарушением разделения центриолей. При этом формируется лишь один полюс, от которого расходятся нити единственного полуверетена. В итоге моноцентрический митоз приводит к полиплоидизации[109].
6) Асимметричный митоз характеризуется непропорциональным развитием противоположных полюсов деления, что приводит к неравномерному распределению хромосом между дочерними ядрами, то есть к анеуплоидии[109]. В результате асимметричный митоз приводит к образованию микроклеток и гигантских клеток с гипо- и гиперплоидными ядрами.
7) Трёхгрупповая метафаза и метафаза с полярными хромосомами характеризуется наличием в метафазе помимо основной экваториальной пластинки ещё двух групп или отдельных («полярных») хромосом в области полюсов деления клетки[109]. Хромосомы сохраняются вблизи полюсов веретена из-за отставания в процессе метакинеза, а не из-за преждевременного расхождения. Причинами отставания могут служить повреждения кинетохора или дезорганизация отдельных хромосомальных нитей, участвующих в движении отстающих хромосом[110].
8) Полая метафаза представляет собой кольцевое скопление хромосом в экваториальной пластинке вдоль периферии клетки[111].
III. Патология митоза, связанная с нарушением цитотомии[править | править код]
Различают две группы патологий митоза, связанные с нарушением цитотомии: раннюю цитотомию, берущую начало ещё в анафазе; либо наоборот, запаздывание или полное отсутствие цитотомии, в результате чего формируются двуядерные клетки, либо образуется одно полиплоидное ядро[111].
Типы митоза[править | править код]
Выработка единой типологии и классификации митозов осложняется целым спектром признаков[~ 9], которые в различных комбинациях создают разнообразие и неоднородность картин митотического деления. При этом отдельные варианты классификации, разработанные применительно к одним таксонам, являются неприемлемыми в отношении других, поскольку не учитывают специфики их митозов. Например, отдельные варианты классификации митозов, свойственных животным или растительным организмам, оказываются неприемлемыми для водорослей[112].
Одним из ключевых признаков, лежащих в основе различных типологий и классификаций митотического деления, является поведение ядерной оболочки. Если образование веретена и само митотическое деление протекает внутри ядра без разрушения ядерной оболочки, то такой тип митоза называют закрытым. Митоз с распадом ядерной оболочки, соответственно, называется открытым, а митоз с распадом оболочки только на полюсах веретена, с образованием «полярных окон» — полузакрытым[112][113].
Также при анализе митоза учитывают особенности веретена деления. При плевромитозе веретено деления состоит из двух полуверетён, располагающихся в метафазе-анафазе под углом друг к другу (также под плевромитозом понимают митоз с боковым по отношению к ядру внеядерным веретеном). Для ортомитоза характерно цельное веретено деления, а в метафазе зачастую наблюдается различимая экваториальная пластинка[113].
В рамках обозначенных признаков наиболее многочисленным является типичный открытый ортомитоз. Этот тип митоза характерен для животных, высших растений и некоторых простейших[114].
Варианты классификации митозов[править | править код]
|
7 типов митоза простейших[113]:
|
6 типов митоза водорослей[112]:
|
 1. закрытый центрический, 2. закрытый ацентрический, 3. полузакрытый центрический, 4. полузакрытый ацентрический, 5. открытый центрический, 6. открытый ацентрический |
Происхождение и эволюция митоза[править | править код]

Предполагается, что сложный митотический процесс высших организмов развивался постепенно из механизмов деления прокариот[115]. Это предположение подтверждается тем, что прокариоты появились приблизительно на миллиард лет раньше первых эукариот. Кроме того, в митозе эукариот и бинарном делении прокариот принимают участие схожие белки.
Возможные промежуточные стадии между бинарным делением и митозом можно проследить у одноклеточных эукариот, у которых в ходе деления не разрушается ядерная мембрана. У большинства же других эукариот, в том числе растений и животных, веретено деления формируется вне ядра, а ядерная оболочка разрушается в течение митоза. Хотя митоз у одноклеточных эукариот ещё недостаточно изучен, можно предположить, что он произошёл от бинарного деления и в конечном счёте достиг того уровня сложности, который имеется у многоклеточных организмов[116].
У многих простейших эукариот митоз также остался процессом, связанным с мембраной, однако теперь уже не плазматической, а ядерной[117]. Возможно, в связи с увеличением размера и числа хромосом, структура типа мезосомы разделилась на два элемента: ЦОМТ на ядерной оболочке и кинетохор на хромосоме. Для соединения этих структур между собой в процессе эволюции развилась промежуточная система микротрубочек. В рамках этого представления, наиболее древним и примитивным считается закрытый внутриядерный плевромитоз. Сегрегация хромосом при этом происходит путём расхождения ЦОМТ, к которым хромосомы крепятся посредством микротрубочек. В свою очередь, ЦОМТ закреплены на ядерной оболочке и расходятся за счёт роста ядерной мембраны между ними[118].
От разных вариантов закрытого внутриядерного плевромитоза, вероятно, берут своё начало несколько параллельных эволюционных линий[118]. В качестве эволюционно прогрессивных признаков при этом рассматриваются: распад ядерной оболочки во время митоза; переход ЦОМТ из ядра в цитоплазму; образование биполярного веретена; усиление спирализации хромосом; формирование экваториальной пластинки в метафазе. Таким образом, эволюция митотического деления идёт в направлении от закрытого внутриядерного плевромитоза к открытому ортомитозу[119].
Эндомитоз[править | править код]
Эндомитоз представляет собой разновидность митоза без деления ядра или клетки, в результате чего в клетке накапливается множество копий одних и тех же хромосом, собранных в одном ядре. Этот процесс может также включать эндоредупликацию, а клетки в этом случае называются эндоплоидными[120]. Примером клеток, подвергающихся эндомитозу, могут служить мегакариоциты, дающие начало тромбоцитам[121].
Крайним случаем эндомитоза является образование гигантских политенных хромосом, появляющихся как результат многократного воспроизведения хромосом без последующего расхождения. Такие хромосомы встречаются в слюнных железах некоторых насекомых, у личинок двукрылых в ядрах клеток кишечника и у некоторых растений в ядрах синергид (например, гороха)[122] .
Значение митоза[править | править код]
Митоз является важным средством поддержания постоянства хромосомного набора. В результате митоза осуществляется идентичное воспроизведение клетки. Следовательно, ключевая роль митоза — копирование генетической информации.
Митоз происходит в следующих случаях:
- Рост и развитие. Количество клеток в организме в процессе роста увеличивается благодаря митозу. Это лежит в развитии многоклеточного организма из единственной клетки — зиготы, а также роста многоклеточного организма.
- Перемещение клеток. В некоторых органах организма, например, коже и пищеварительном тракте, клетки постоянно отшелушиваются и заменяются новыми. Новые клетки образуются путём митоза, а потому являются точными копиями своих предшественников. Схожим путём происходит замена красных кровяных клеток — эритроцитов, имеющих короткую продолжительность жизни — 4 месяца, а новые эритроциты формируются путём митоза.
- Регенерация. Некоторые организмы способны восстанавливать утраченные части тела. В этих случаях образование новых клеток часто идёт путём митоза. Например, благодаря митозу морские звёзды восстанавливают утраченные лучи.
- Бесполое размножение. Некоторые организмы образуют генетически идентичное потомство путём бесполого размножения. Например, гидры размножаются бесполым способом при помощи почкования. Поверхностные клетки гидры подвергаются митозу и образуют скопления клеток, называемые почками. Митоз продолжается и в клетках почки, и она вырастает во взрослую особь. Сходное клеточное деление происходит при вегетативном размножении растений.
См. также[править | править код]
Примечания[править | править код]
- Комментарии
- ↑ Сам факт разделения морфологии митотического веретена на два типа не отменяет возможности сочетания обоих в пределах одного организма. Например, в раннем эмбриогенезе млекопитающих при делении созревания ооцита и при I и II делении зиготы наблюдаются бесцентриолярные анастральные митозы. Но уже начиная с третьего клеточного деления и во всех последующих, клетки делятся при участии астральных веретен, в полюсах которых всегда обнаруживаются центриоли[29]
- ↑ Первоначально, исходя из морфологических особенностей митоза, этот процесс был разделен лишь на четыре основные стадии: профазу, метафазу, анафазу и телофазу[38].
- ↑ Если митотические клетки поместить в тяжелую воду (D2O) или обработать таксолом (эти воздействия подавляют разборку микротрубочек), то нити веретена будут удлиняться. Такое стабилизированное веретено не может тянуть хромосомы, и митоз останавливается. Но митоз блокируется и при прямо противоположном воздействии, если нити веретена обратимо разрушить с помощью одного из трех агентов, подавляющих сборку тубулина в микротрубочки, – колхицина, низкой температуры или высокого гидростатического давления[49].
- ↑ Существует, как минимум, три гипотетические модели, объясняющие вероятный механизм расхождения хромосом в анафазе А. По одной из них перемещение хроматид объясняется наличием в кинетохоре «шагающих» белков, сходных по своей природе с динеином или кинезином; они продвигаются по микротрубочке, используя для этого энергию гидролиза АТФ. По другой гипотезе движение хромосом обусловлено распадом микротрубочек: по мере того как субъединицы тубулина диссоциируют, кинетохор, чтобы сохранить связь с микротрубочкой, должен скользить в направлении полюса. Третья возможность состоит в том, что микротрубочки не ответственны прямо за возникновение силы, движущей кинетохор к полюсам, а просто регулируют движение, вызываемое какой-то другой структурой[63].
- ↑ Существует целый ряд примеров, описывающих многократные митотические деления ядер без сопутствующего разделения клеточного тела. Так, в эндосперме многих растений протекают множественные митозы без деления цитоплазмы, что приводит к образованию многоядерного симпласта. Аналогичная ситуация наблюдается при синхронных делениях многочисленных ядер миксомицетов, или на ранних этапах развития зародышей некоторых насекомых[70].
- ↑ В качестве возможных рассматриваются несколько инициирующих моделей. Например, предполагается, что комплексы M-Cdk1 не полностью блокируются семейством ингибиторов Wee1. В результате, пропорционально увеличению концентрации митотических циклинов, к началу профазы может накопиться критическая масса активных киназ M-Cdk1. Частичная активация циклин-киназ у позвоночных, возможно, обеспечивается фосфатазой Cdc25B, уровень активности которой возрастает с поздней S-фазы и достигает максимума в профазе митоза. Однако было продемонстрировано, что мышиные клетки способны делиться в отсутствие этого стимула. Ещё одним возможным активатором может быть комплекс циклин A-Cdk, сохраняющий свою активность с начала S-фазы вплоть до конца прометафазы митоза[85].
- ↑ После инфицирования культур диплоидных клеток лёгкого человека вирусом Herpes simplex число патологических митозов (к-митозы, хромосомные аберрации) увеличивалось с 3 % в контроле до 40—60 % в инфицированной культуре[97].
- ↑ На примере эпителия гортани человека были получены данные об увеличении числа патологических митозов при раке. Если при хроническом воспалении и в папилломах «юношеского» типа количество патологических митозов всего в 2—2,5 раза превышало их число в нормальном эпителии, то при предраке количество патологических митозов составляло около 25 %, а при раке и атипическом папилломатозе с переходом в рак оно достигало 36—45 %[98].
- ↑ К таким признакам, например, относятся: поведение ядерной оболочки со всеми переходами от интактной, в разной степени фрагментированной до полностью распадающейся; неоднозначное поведение ядрышка от сохраняющегося до частично или полностью исчезающего; разная степень спирализации (или полное отсутствие таковой у динофлагеллат) и морфологической дифференциации хромосом; особенности расположения хромосом в метафазной пластинке; наличие кинетохоров и различия в их организации; различия в морфологии, характере заложения и организации веретена, продолжительность сохранения его межзональной зоны; появление наряду с центриолями особых полярных образований различной организации и места своей локализации; разная степень развития перинуклеарной оболочки и т. п.[112]
- Источники
- ↑ Большая Медицинская Энциклопедия (БМЭ), под редакцией Петровского Б. В., 3-е издание, том 10 / Кариокинез
- ↑ Кариокинез // Большая советская энциклопедия : [в 51 т.] / гл. ред. С. И. Вавилов. — 2-е изд. — М. : Советская энциклопедия, 1949—1958.
- ↑ 1 2 3 4 Биологический энциклопедический словарь / Гл. редактор Гиляров М. С.. — М.: Сов. энциклопедия, 1986. — 831 с. — 100 000 экз.
- ↑ Алов, 1974, с. 335.
- ↑ Гилберт, 1995, с. 202.
- ↑ 1 2 Албертс и др., 1993, с. 396.
- ↑ 1 2 Алов, 1974, с. 336.
- ↑ Булдаков, Калистратова, 2003, с. 39.
- ↑ Raikov, I. B. The diversity of forms of mitosis in protozoa: A comparative review (англ.) // European Journal of Protistology : journal. — 1994. — Vol. 30, no. 3. — P. 253—269. — doi:10.1016/S0932-4739(11)80072-6.
- ↑ De Souza C. P., Osmani S. A. Mitosis, not just open or closed (неопр.) // Eukaryotic Cell. — 2007. — September (т. 6, № 9). — С. 1521—1527. — doi:10.1128/EC.00178-07. — PMID 17660363. — PMC 2043359.
- ↑ Robert C. King. A dictionary of genetics. — 7th ed. — Oxford: Oxford University Press, 2006. — x, 596 pages с. — ISBN 978-0-19-530762-7, 0-19-530762-3, 978-0-19-530761-0, 0-19-530761-5, 0-19-514325-6, 978-0-19-514325-6, 978-0-19-513076-8, 0-19-513076-6, 978-0-19-989142-9, 0-19-989142-7.
- ↑ 1 2 История биологии до начала XX века, 1972, с. 489.
- ↑ История биологии до начала XX века, 1972, с. 485.
- ↑ Gloria Robinson. Schneider, Friedrich Anton (англ.). www.encyclopedia.com. Дата обращения: 25 апреля 2017. Архивировано 26 апреля 2017 года.
- ↑ История биологии до начала XX века, 1972, с. 486.
- ↑ История биологии до начала XX века, 1972, с. 487.
- ↑ История биологии до начала XX века, 1972, с. 488.
- ↑ 1 2 История биологии до начала XX века, 1972, с. 491.
- ↑ Ченцов, 2004, с. 470.
- ↑ Албертс и др., 1993, с. 400.
- ↑ Ченцов, 2004, с. 471.
- ↑ Албертс и др., 1993, с. 403.
- ↑ Албертс и др., 1993, с. 404—405.
- ↑ Албертс и др., 1993, с. 438.
- ↑ Албертс и др., 1993, с. 463.
- ↑ NIGMS — From Molecules to Medicines: Cell Biology and Biophysics (англ.). Архивировано из оригинала 11 февраля 2012 года.
- ↑ 1 2 Албертс и др., 1993, с. 439.
- ↑ 1 2 3 4 5 Албертс и др., 1993, с. 444.
- ↑ Албертс и др., 1993, с. 429.
- ↑ 1 2 3 Ченцов, 2004, с. 429.
- ↑ Ченцов, 2004, с. 430.
- ↑ 1 2 Албертс и др., 1993, с. 446.
- ↑ 1 2 Ченцов, 2004, с. 433.
- ↑ 1 2 Ченцов, 2004, с. 434.
- ↑ Алов, 1972, с. 18.
- ↑ Албертс и др., 1993, с. 415.
- ↑ Алов, 1972, с. 21.
- ↑ Алов, 1972, с. 12.
- ↑ Алов, 1972, с. 19.
- ↑ 1 2 3 4 Lackie, 2013, p. 531.
- ↑ Evert, Eichhorn, 2013, p. 65.
- ↑ Льюин и др., 2011, с. 876.
- ↑ Smith L. G. Division Plane Determination in Plant Cells (англ.) (май 2006). Архивировано 25 февраля 2015 года.
- ↑ Льюин и др., 2011, с. 932.
- ↑ 1 2 3 Льюин и др., 2011, с. 877.
- ↑ 1 2 3 4 5 6 Morgan, 2007, p. 89.
- ↑ Алов, 1972, с. 83.
- ↑ 1 2 3 Ченцов, 2004, с. 434.
- ↑ 1 2 Албертс и др., 1993, с. 445.
- ↑ Ченцов, 2004, с. 436.
- ↑ Ченцов, 2004, с. 436—437.
- ↑ 1 2 3 Ченцов, 2004, с. 436.
- ↑ 1 2 Албертс и др., 1993, с. 448.
- ↑ Албертс и др., 1993, с. 449.
- ↑ Алов, 1972, с. 108.
- ↑ Алов, 1972, с. 112.
- ↑ 1 2 3 Ченцов, 2004, с. 439.
- ↑ Алов, 1972, с. 113.
- ↑ Албертс и др., 1993, с. 451.
- ↑ 1 2 Албертс и др., 1993, с. 452.
- ↑ Ченцов, 2004, с. 440.
- ↑ Алов, 1972, с. 119.
- ↑ 1 2 3 4 Албертс и др., 1993, с. 453.
- ↑ Ченцов, 2004, с. 441.
- ↑ Албертс и др., 1993, с. 454.
- ↑ 1 2 3 Ченцов, 2004, с. 442.
- ↑ Алов, 1972, с. 135.
- ↑ Албертс и др., 1993, с. 457.
- ↑ Алов, 1972, с. 137.
- ↑ 1 2 Албертс и др., 1993, с. 458.
- ↑ Алов, 1972, с. 140.
- ↑ Албертс и др., 1993, с. 459.
- ↑ Албертс и др., 1993, с. 460.
- ↑ Албертс и др., 1993, с. 461.
- ↑ Morgan, 2007, p. 90.
- ↑ 1 2 Morgan, 2007, p. 91.
- ↑ Alberts at al., 2008, p. 1071.
- ↑ Morgan, 2007, p. 99.
- ↑ Morgan, 2007, p. 30.
- ↑ Morgan, 2007, p. 28.
- ↑ 1 2 Morgan, 2007, p. 32.
- ↑ Morgan, 2007, p. 97.
- ↑ 1 2 3 Alberts at al., 2008, p. 1074.
- ↑ Morgan, 2007, p. 96.
- ↑ Morgan, 2007, p. 98—99.
- ↑ Morgan, 2007, p. 98.
- ↑ Morgan, 2007, p. 102.
- ↑ Morgan, 2007, p. 103.
- ↑ 1 2 3 4 Morgan, 2007, p. 48.
- ↑ Morgan, 2007, p. 46.
- ↑ Alberts at al., 2008, p. 1087.
- ↑ Lackie, 2013, p. 419.
- ↑ Redei, 2008, p. 1238—1239.
- ↑ 1 2 Hartwell et al., 2010, p. 146.
- ↑ 1 2 3 Redei, 2008, p. 1239.
- ↑ Redei, 2008, p. 1240.
- ↑ Алов, 1972, с. 192.
- ↑ Алов, 1972, с. 193.
- ↑ 1 2 Алов, 1972, с. 167.
- ↑ 1 2 3 Алов, 1972, с. 169.
- ↑ Алов, 1972, с. 170.
- ↑ Алов, 1972, с. 171.
- ↑ 1 2 Алов, 1972, с. 172.
- ↑ 1 2 Алов, 1972, с. 174.
- ↑ 1 2 Алов, 1972, с. 176.
- ↑ Алов, 1972, с. 177.
- ↑ 1 2 Алов, 1972, с. 183.
- ↑ Алов, 1972, с. 184.
- ↑ 1 2 3 Алов, 1972, с. 185.
- ↑ Алов, 1972, с. 186.
- ↑ 1 2 Алов, 1972, с. 188.
- ↑ 1 2 3 4 Седова, 1996, с. 103.
- ↑ 1 2 3 Райков, 1978, с. 57.
- ↑ Ченцов, 2004, с. 428.
- ↑ Албертс и др., 1993, с. 465.
- ↑ The Mitotic Phase and the G0 Phase (англ.). Boundless. Дата обращения: 25 апреля 2017. Архивировано из оригинала 26 апреля 2017 года.
- ↑ Райков, 1978, с. 93.
- ↑ 1 2 Райков, 1978, с. 94.
- ↑ Райков, 1978, с. 95.
- ↑ Lilly M., Duronio R. New insights into cell cycle control from the Drosophila endocycle (англ.) // Oncogene : journal. — 2005. — Vol. 24, no. 17. — P. 2765—2775. — doi:10.1038/sj.onc.1208610. — PMID 15838513.
- ↑ Italiano J. E., Shivdasani RA. Megakaryocytes and beyond: the birth of platelets (англ.) // Journal of Thrombosis and Haemostasis : journal. — 2003. — Vol. 1, no. 6. — P. 1174—1182. — doi:10.1046/j.1538-7836.2003.00290.x. — PMID 12871316.
- ↑ Инге-Вечтомов, 2010, с. 89—90.
Литература[править | править код]
- Alberts B. at al. Molecular biology of the cell. — 5 edition. — Garland science, 2008. — 1601 p. — ISBN 978-0-8153-4105.
- Lackie J. M. (ed.). The dictionary of cell and molecular biology. — 5 edition. — Academic Press, 2013. — 750 p. — ISBN 978-0-12-384931-1.
- Hartwell L. et al. Genetics: from genes to genomes. — 4 edition. — McGraw-Hill Science, 2010. — 816 p. — ISBN 978–0–07–352526–6.
- Morgan D. O. The cell cycle: principles of control. — New science press, 2007. — 297 p. — ISBN 978-0-9539181-2-6.
- Evert R. F., Eichhorn S. E. Raven biology of plants. — 8 edition. — W. H. Freeman and Company, 2013. — 880 p. — ISBN 978-1-4292-1961-7.
- Redei G. P. (ed.). Encyclopedia of genetics, genomics, proteomics, and informatics. — 3 edition. — Springer, 2008. — 1822 p. — ISBN 978-1-4020-6753-2.
- Алов И. А. Цитофизиология и патология митоза. — М.: «Медицина», 1972. — 264 с. — 3700 экз.
- Митоз / И. А. Алов // Большая Советская Энциклопедия / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская Энциклопедия, 1974. — Т. 16 : Мёзия — Моршанск. — С. 335—337. — 629 000 экз.
- Албертс Б. и др. Молекулярная биология клетки: В 3 т. — 2-е изд., перераб. — М.: «Мир», 1993. — Т. 2. — 539 с. — ISBN 5-03-001987-1.
- Биологический энциклопедический словарь / Гл. редактор Гиляров М. С.. — М.: «Советская энциклопедия», 1986. — 831 с. — 100 000 экз.
- Булдаков Л. А., Калистратова В. С. Радиоактивное излучение и здоровье. — М.: Информ-Атом, 2003. — 165 с. Архивировано 2 декабря 2003 года.
- Гилберт С. Биология развития: в 3 томах. — М.: «Мир», 1995. — Т. 3. — 352 с. — 5000 экз. — ISBN 5-03-001833-6.
- История биологии с древнейших времён до начала XX века / Под редакцией С. Р. Микулинского. — М.: «Наука», 1972. — 564 с. — 9600 экз.
- Льюин Б. и др. Клетки. — М.: БИНОМ. Лаборатория знаний, 2011. — 951 с. — (Лучший зарубежный учебник). — ISBN 978-5-94774-794-2.
- Мэзия Д. Митоз и физиология клеточного деления = D. Mazia. Mitosis and the physiology of cell division. The cell. Ed. by J. Brachet and A. Mirsky. Vol. III. New York-London, Acad. press, 1961 / Пер. с англ. Д. М. Кершнера; Под. ред. и с предисл. проф. Л. Н. Жинкина. — М.: Изд-во иностр. лит., 1963. — 428, [64] с.
- Райков И. Б. Ядро простейших. Морфология и эволюция. — Л.: «Наука», 1978. — 328 с. — 1600 экз.
- Седова Т. В. Кариология водорослей. — СПб.: «Наука», 1996. — 386 с. — 500 экз. — ISBN 5-02-026058-4.
- Ченцов Ю. С. Введение в клеточную биологию: Учебник для вузов. — 4-е изд., переработанное и дополненное. — М.: ИКЦ «Академкнига», 2004. — 495 с. — 3000 экз. — ISBN 5-94628-105-4.
- Инге-Вечтомов С. Г. Генетика с основами селекции. — 2-е изд., переработанное и дополненное. — СПб.: Издательство Н-Л, 2010. — 718 с. — 3000 экз. — ISBN 978-5-94869-105-3.
Ссылки[править | править код]
Иллюстрации[править | править код]
- Тематическая выборка иллюстраций в базе Science Photo Library (англ.). www.sciencephoto.com. Архивировано 5 августа 2012 года.
Анимация[править | править код]
- Стадии митоза (анимированное видео) (англ.) (18 октября 2009).
- Стадии митоза (трёхмерная анимация № 1) (англ.). Hybrid Medical Animation (15 июня 2009).
- Стадии митоза (трёхмерная анимация № 2) (англ.). Protein Lounge (20 октября 2009).
Видео[править | править код]
- Тематическая выборка видеороликов и анимации в базе Science Photo Library (англ.). www.sciencephoto.com. — Для просмотра необходим QuickTime Player. Архивировано 5 августа 2012 года.
- Виллиам Салливан, Клаудио Санкел и др. Синхронные митотические деления (видео № 1) (англ.). Molecular Biology of the Cell, 5th Edition (Media DVD-ROM). Garland Science (2002).
- Синхронные митотические деления (видео № 2) (25 ноября 2011).
- Митотическое деление животной клетки (видео № 1) (27 мая 2009).
- Митотическое деление животной клетки (видео № 2) (29 марта 2010).
- Митотическое деление растительной клетки (видео № 1) (6 мая 2007).
- Митотическое деление растительной клетки (видео № 2, начиная с 0:58) (англ.) (20 июля 2010).
| В библиографических каталогах |
|---|
| Фазы |
|  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Регуляторы |
| ||||||||||
| Основное |  | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Классификация | |||||||||||
| Структура |
| ||||||||||
| Перестройки и нарушения | |||||||||||
| Хромосомное определение пола | |||||||||||
| Методы | |||||||||||